Investigation into the long-finned pilot whale (Globicephala melas) mass stranding event, Tolsta, na h-Eileanan Siar, July 2023
This report details the comprehensive analysis into the mass stranding event (MSE) of long-finned pilot whales that occurred in Tolsta Bay, Isle of Lewis, Scotland, on 16 July 2023.
Section 7: Postmortem (Necropsy) Examinations
Diagnostic necropsies form the foundation of marine mammal stranding investigations. Their primary objective is to evaluate individual health and identify factors contributing to stranding, with a secondary objective of collecting samples for further analyses. These samples are used to investigate causes of morbidity and mortality at the individual level and to assess population-level metrics of ecology, physiology, and health, including diet, contaminant exposure, and life-history parameters. Necropsies are often conducted under significant time and logistical constraints, particularly when working with large carcasses outside dedicated facilities, necessitating rapid recovery of data and samples before decomposition degrades information quality. All examinations followed internationally agreed protocols (Ijsseldijk et al., 2019).
A comprehensive suite of tissues and measurements were collected to support diagnostic assessment and downstream analyses. Such systematic tissue sampling is essential for identifying acute pathological processes and chronic health stressors, while also contributing to broader understanding of population dynamics, foraging ecology, vital rates, and anthropogenic impacts. Selected tissues were archived to enable future investigations as new research questions arise.
7.1 Sample Collection
All animals had the following data recorded and samples taken:
- Detailed morphometrics including total length, girth (both in front of the dorsal fin, and behind the pectoral fins), and blowhole-to-dorsal fin measurements.
- Photographs.
- Age class determined.
- External examination for trauma, bycatch or entanglement marks.
- Blubber thickness.
- Sex and, where possible, reproductive status.
- Stomach contents and evidence of recent feeding.
- Samples of skin, blubber, muscle, and liver.
- Teeth (for ageing)
Most of the animals which were examined at necropsy also had the following additional data recorded and samples taken:
- Weight.
- Bacteriology (lung, liver, kidney, brain, spleen as minimum).
- Formalin-fixed samples for histopathology.
- Body condition scoring (good/moderate/poor).
- Cochlea (eight animals).
- Urine and faeces.
7.2 Health assessment parameters
In addition to the samples outlined above, the following assessments were made on all animals taken to necropsy:
- Bruising patterns (subcutaneous, sub-mandibular)- an indicator of live stranding.
- Lung asymmetry detection.
- Water/sand aspiration and ingestion.
- Evidence of hepatic lipidosis (fatty liver).
- Parasite burden, including subcutaneous cysts/abscesses (scored for severity).
- Reproductive status assessment (if pregnant or evidence of recent parturition). This was also done for animals not necropsied.
- Ovary evaluation inclusion.
- Foetal size measurement.
- Lactation status.
All carcasses were sampled (skin, blubber, and muscle) the day after stranding prior to being transported to Stornoway landfill site for necropsy. (Figure 10 to Figure 12). Between 17 and 21 July, SMASS conducted full necropsies on 23 individuals, representing 43% of the group. This was the maximum feasible number before decomposition compromised diagnostic quality. The remaining 31 cases (57%) were photographed, measured, and sampled for liver, muscle, and where possible, brain tissue to enable toxicological analysis, age determination, and reproductive assessment. Cases were prioritised for full necropsy based on decomposition status, targeting the freshest individuals while maintaining representative age and sex distributions.
All animals were evaluated for external indicators of poor body condition, disease, or pre-existing injury that might suggest a sick or compromised individual could have led otherwise healthy animals ashore.



7.3 Necropsy Findings
All necropsied animals were in good nutritional condition with adequate blubber reserves. External body shape, blubber thickness, and back muscle mass were within normal ranges, and free lipid was evident on cut section in all cases, indicating sufficient energy stores. No animals showed evidence of significant or prolonged negative energy balance; in particular, there were no metabolic hepatic lesions such as jaundice or fatty liver.
Of the 23 animals examined, ten had been euthanised. Among the remaining 13 animals, the proximal cause of death in ten cases was either drowning or pathology consistent with hyperthermia associated with the stranding process. Drowning was diagnosed by the presence of sand or silt in the airways, consistent with aspiration of seawater and sediment, most likely in the surf zone. Stranding-related pathology was identified by lung asymmetry, deep tissue bruising, and/or myoglobinuria. These findings strongly indicate that the animals were alive at the time of stranding but succumbed either during the initial beaching or shortly thereafter.
Twelve animals exhibited chronic subcuticular parasitic infections, evident as multifocal, well-demarcated abscesses up to 30 cm in diameter containing white purulent material. These lesions were likely of parasitic origin, with possible secondary bacterial infection, but showed no evidence of dissemination, sepsis, or systemic compromise. All animals were in good nutritional condition with adequate blubber reserves and no indication of infectious disease, significant trauma, fisheries interaction, or proximity to underwater explosions. Stomachs contained seawater but no recent ingesta, suggesting regurgitation prior to death. In non-euthanised animals, drowning due to aspiration of seawater and sand on the shallow beach was the primary cause of death. Euthanasia by ballistic trauma (n = 12) was effective, producing extensive cranial injury consistent with instantaneous death.
One female (M371.40/23), initially suspected to have a uterine prolapse, was found to have prolapsed placental tissue, consistent with ongoing, possibly prolonged parturition. The cervix was dilated, and a full-term male foetus in normal (tail-first) presentation occupied the left uterine horn. Although no physical obstruction to delivery was present, placental separation, pulmonary oedema, and meconium expulsion indicated foetal distress, suggesting dystocia. The placental tissue appeared otherwise normal, and the foetus was likely alive at or near the time of maternal death, implying that the stranding event may have precipitated or exacerbated parturition difficulties. No other animals showed evidence of acute disease or injury on gross examination, though undetected pathology in non-necropsied cases cannot be excluded.
7.4 Reproductive Status
Several females were noted to be pregnant in various stages of gestation. Estimated, from foetal size, females had either recently conceived (in the past few weeks) or were pregnant with at-term foetuses. In the latter case there were two animals with prolapsed placenta from the right (non-gravid) horn during the stranding process. There were additionally nine females showing evidence of recent parturition. These findings align with a gestation time estimated to be approximately 12 months. Given the strong matrilineal, intergenerational bonds in this species, it is possible the group had clustered around one or more females who were in the process of giving birth and this may have contributed to so many animals coming ashore.
7.5 Fisheries Interactions
There was no evidence of recent or historic trauma to any of these animals which may be attributed to bycatch, entanglement or ingestion of significant marine debris. This included no indication of either recent or historical skin lesions or scarring consistent in pattern to those caused by either net or line entanglement or entrapment. One animal had a small loop of green braided polyethylene twine in the stomach consistent with a type widely used in fishing and some land-based industries, but this was not causing problems and was considered an incidental finding.
7.6 Microbiological Analysis
Comprehensive microbiological and histological examinations were undertaken on all necropsied cases to assess pathogen carriage, prevalence, and potential subclinical infection as a component of evaluating overall health status. This investigation included microbiological culture of at least five organs (brain, lung, spleen, liver, and kidney), histological examination, and viral PCR screening for morbillivirus. High pathogenicity avian influenza virus (HPAIV) was not suspected in any of the cases based on clinical presentation or gross pathological findings.
7.6.1 Bacteriology Screening
Nick Davison and the Veterinary Diagnostic Services (VDS); University of Glasgow.
Samples from various organs collected during necropsy were stored frozen until screened by standard microbiological culture techniques for any potential bacterial infections likely to be implicated in the mass stranding event. Given the ‘sick leader’ hypothesis has often been raised as potential cause for these events and a bacterial infection of the central nervous system (CNS) would not always be evident on gross necropsy and hence bacterial culture was indicated. Additionally, given the observed subcutaneous abscesses in a number of cases, the investigation sought to ascertain if any animals showed indication of pathogenic bacterial infection or disseminated bacteriaemia or sepsis.
Tissues from 23 animals were subjected to bacteriological examination (15 adults, seven juveniles and one calf). Isolates were recovered from all cases subjected to culture. Routinely lung, liver, spleen, brain and when available CSF (cerebrospinal fluid) cultures as standard. Molecular techniques were not used.
Bacteriological culture was carried out as per Foster et al (2002). Briefly, tissue samples were inoculated directly onto Columbia sheep blood agar (CSBA) (Oxoid) and Farrell’s medium (Animal and Plant Health Agency), incubated at 37°C in a capnophilic (5% CO2) atmosphere and examined daily for 14 days. Samples were also inoculated directly onto MacConkey agar (Oxoid) and incubated at 37°C in air. This investigation included specialised cultures for Brucella species given the known association with brucella infection and meningoencephalitis in cetaceans, including long-finned pilot whales, which has been implicated in live strandings (Davison et al. 2015). Lung was cultured in all cases. Liver was cultured in 22 cases. Spleen was cultured in 21 cases. Brain was cultured in 13 cases and CSF was available for culture in four animals. Lesions with purulent material were observed on 12 animals and cultured in five cases.
Results: Most recovered isolates were Edwardsiella tarda and Enterococcus faecalis followed by E. coli, Aeromonas sp., and Vibrio sp. These isolates were considered to be incidental and are likely postmortem invaders. A single case involving Brucella ceti was isolated in mixed culture from the purulent abscess caudal/cranial to abdominal in M371.43/23 (Figure 13). The significance of this isolate is uncertain, but in the absence of associated pathology it was considered an incidental finding. The other abscesses cultured either proved sterile or revealed postmortem invaders such as Enterococcus sp., Edwardsiella sp., and E. coli.
7.6.2 Viral Screening
Madeleine Maley & Mara Rocchi, Moredun Research Institute.
Cetacean morbillivirus (CeMV), the etiological agent of distemper in cetaceans, is a significant cause of morbidity and mortality globally and has been associated with several epizootic events (Sierra et al., 2020; Van Bressem et al., 2008).
To investigate the potential involvement of CeMV involvement in the MSE, total RNA was extracted from frozen (-80 °C) lung and pooled tissue samples (n = 23). Reverse transcriptase polymerase chain reaction (RT-PCR) assays targeting the conserved N-terminal region of the morbillivirus nucleoprotein (N) gene were performed in duplicate (Grant et al. 2009).
Results: No cetacean morbillivirus CeMV RNA was detected. In addition, necropsy findings revealed no macroscopic or histological lesions consistent with morbilliviral infection. Consequently, cetacean morbillivirus was excluded as a differential in this stranding event.
7.7 Histopathology
Mark Dagleish and the Veterinary Diagnostic Services (VDS), University of Glasgow.
Histological analysis of stranded pilot whales was constrained by tissue autolysis and only 14 of the 18 cases examined histologically were considered diagnostic.
Results: Histological examinations revealed a range of pathological lesions commonly encountered in cetacean standings. These included mild tissue swelling (oedema) of brain, lung and heart tissue, and brain and lung haemorrhages consistent with live stranding and associated hyperthermia. There was also evidence of liver changes including fatty deposits and iron accumulation, muscle degeneration, localised inflammatory skin conditions, and parasitic infections affecting the stomach and mammary glands. However, the extent and severity of these lesions were insufficient to be considered a primary cause of morbidity or mortality. Overall, the group appeared to be in good health with no major underlying diseases identified. This supports the notion that the animals were healthy prior to stranding.
7.8 Teeth aging
Dr Ailsa Hall, Prof Simon Northridge and Simon Moss; Sea Mammal Research Unit, University of St Andrews.
A modified acid etching method based on Pierce and Kaijmura (1980) and Martin et al. (1987) was used to investigate age-length relationships. Teeth were bisected longitudinally and a 0.5mm central section was etched in 5% formic acid for 1-2 hours. Growth layers were counted from digital images taken under light microscopy at 2.5x magnification and enhanced using microscope camera software or Adobe Photoshop where necessary.
A subset of 16 teeth were independently counted by a second reader, with ages differing by a maximum of ±4 years (average difference 16%, or 3% excluding two worn teeth from older animals). Teeth were also weighed as an additional parameter for estimating age and body size. Gompertz growth curves were fitted to total length (tip of upper jaw to tail notch) and age data.
Results: A significant linear relationship between tooth mass and age was found (p<0.001) but low R2 values (0.40 and 0.55 for females and males respectively) suggest tooth mass is not a good predictor but might be used as a rough guide
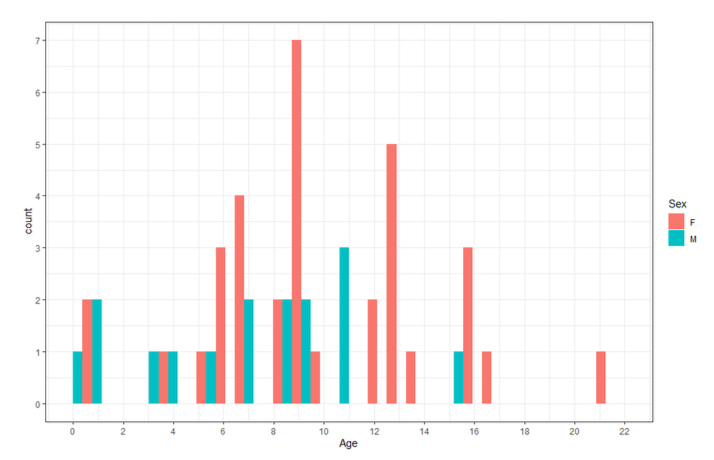
The distribution of ages in the sample for males and females is shown in Figure 14 with a mean age of 10 +/- 5 years for females and 7 +/- 4 years for males.
Previous studies had found that maximum length for the males was 630 cm and 546 cm for the females with the oldest male in the sample being 20 years and the oldest female being 25 years. Mature females were generally > 400 cm and older than 12 years. The data from this MSE corresponds to these results, with mature females (as determined from their reproductive tracts) showing sexual maturity around 400 cm in length and at a minimum age of 10 years. The minimum age for mature males in this group was 9 years old.
Contact
Email: marine_species@gov.scot