Offshore wind developments - collision and displacement in petrels and shearwaters: literature review
Literature review of the risk of collision and displacement in petrels and shearwaters from offshore wind developments in Scotland.
4 Species accounts
4.1 General ecology
The five seabird species that are the subject of this review all belong to the Order Procellariiformes, which includes the most pelagic of all seabirds. The three species which are the primary focus (Manx Shearwater, European Storm-petrel and Leach's Storm-petrel) are small-bodied and relative immobile on land, which renders them vulnerable to predation, so breeding colonies are restricted to islands that are often very remote, where the birds nest out of sight in burrows and cavities, and only attend the colony at night. The breeding season is divided into four distinct stages. During the pre-laying stage birds occupy and defend nest crevices, attract a mate and copulate. Females typically spend a protracted period (c. one week) at sea prior to laying (the so-called "pre-laying exodus") to acquire the nutrients needed to produce the single large egg. Once laid, the egg is incubated for around six weeks (the "incubation period"), until hatching. The newly hatched chick is incapable of full thermoregulation and parents take turns brooding the chick at the nest whilst the other feeds at sea ("brooding period"). Once chicks acquire the ability to thermoregulate independently, both adults spend the day feeding at sea, returning during darkness at intervals of 1 to 7 days to feed the chick (the "post-brooding" period). Although the storm-petrels are little bigger than a sparrow, and shearwaters the size of a pigeon, these species may cover thousands of kilometres at sea on a single foraging trip and the maximum range of a trip may lie hundreds of kilometres from the colony.
The eggs and chicks have very protracted development, requiring nearly four months from egg laying to fledging. For European Storm-petrels, within a particular colony the timing of breeding may be poorly synchronised among individuals, such that there will be birds engaged in breeding activities for more than six months of the year. The combination of large foraging ranges and very protracted breeding seasons means that birds will be exposed to risks from marine activities over a wider geographic area, and for a longer period of the year, than many other seabird species. All three focal species are trans-equatorial migrants, and immature birds do not return to the breeding colonies for several years, presumably remaining within, or close to, the wintering grounds in the South Atlantic. Storm-petrels and shearwaters do not reach breeding age for 3–6 years which, coupled with low fecundity, means that adults need to maintain a high annual survival rate for population stability, and any adverse impact on adult survival will have large, and long-term, consequences for population size.
While most of the above also applies to the two additional species included in this review, Northern Fulmars, and sometimes Sooty Shearwaters, will nest on mainland coasts as well as islands. Unlike the other species reviewed here, the Northern Fulmar is a surface nester, will attend its nest both during the day and at night, and is present in Scottish waters during both its breeding and non-breeding periods.
4.1.1 Note on spatial and temporal distribution
Information on the distribution of seabirds at sea is available from boat and aerial surveys, and more recently from tracking birds during the breeding season, and during their migration and non-breeding seasons. Both methods suffer from various drawbacks: aerial and vessel-based surveys tend to under-represent storm-petrel occurrence/density due to the birds' small size and dark colouration, and at-sea surveys are unable to determine the provenance, age or breeding stage of birds encountered at sea. The European Seabirds At Sea (ESAS) database used to produce maps of marine distribution of seabird (e.g. Stone et al., 1994, Waggitt et al., 2020) includes data collected over many decades and may not reliably reflect current marine distribution of seabirds, due to changing seabird population sizes, breeding colony distributions and prey distribution in recent decades. Tracking studies usually provide data on a relatively small number of birds from a given colony in a particular year and GPS tracking is almost exclusively restricted to the breeding season as birds must be recaptured to retrieve the tags and download the data. Therefore, tracking studies tend to provide a snapshot of the movements of a small number of birds within a restricted period. This snapshot may not always be representative of the foraging movements of the wider colony, particularly as the tagging itself may influence the behaviour of the birds (e.g.Gillies et al., 2020). Geolocators (GLS) can be used to track year-round movements but at a much lower resolution (two locations per day) and, again, the tags must be retrieved to download the data. The necessity of tag retrieval means that tracking studies for these species focus almost exclusively on breeding adults that return reliably to the colony, but tracking of juveniles and immature birds, which may not return to their natal colony for several years, presents methodological challenges.
4.1.2 Note on population status and abundance
The most complete population estimates available for seabirds breeding in Scotland are currently from Mitchell et al. (2004), which summarises the results of Seabird 2000, a census of Britain and Ireland's breeding seabirds, conducted between 1998 and 2002. The results of the latest census, Seabirds Count conducted between…, are due to be published in 2023 and will provide an important update on Scotland's seabird population status and trends.
4.2 Manx Shearwater
4.2.1 Spatial and temporal distribution in Scotland
4.2.1.1 Breeding distribution
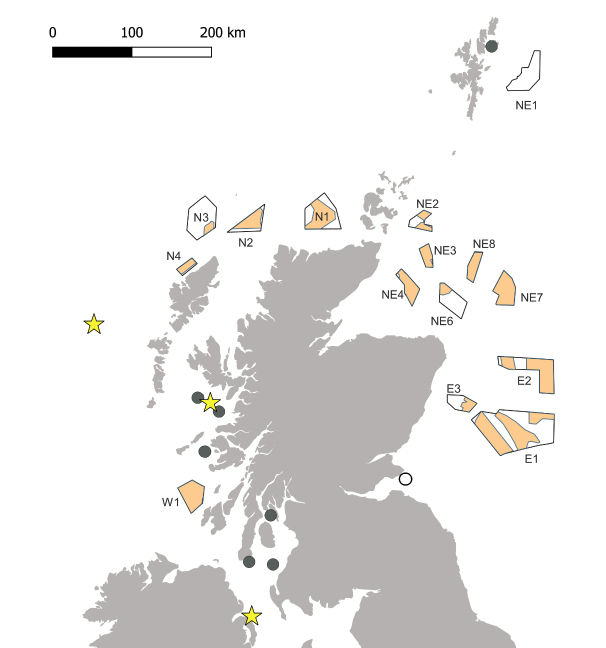
The UK and Ireland's breeding colonies of Manx Shearwater are all located on islands, almost entirely on the western fringes, with additional colonies on the Irish Sea coasts (Figure 1). Very small numbers (< 10 pairs) formerly bred in Orkney (Cramp et al., 1974) and Shetland (Mitchell et al., 2004) but recent confirmation of breeding in these archipelagos is lacking. The species is present in Scottish waters between April and October, with nests occupied between April and August (Harris, 1966b, Guilford et al., 2009, Waggitt et al., 2020).
4.2.1.2 Marine distribution
4.2.1.2.1 Derived from at-sea surveys
Observations from offshore surveys in north-west European waters between 1979 and 1993 are collated in Stone et al. (1994) and show that, during the breeding season, the highest concentrations of birds are around the largest colonies of Rum (west Scotland), and Skomer and Skokholm (south-west Wales). During April, the highest densities were in the inshore waters of west Scotland. Feeding birds were observed in the offshore waters of west Scotland in May and June but occurred in inshore waters later in the season, during chick-rearing, when high densities of birds were present. This change in distribution is likely explained by the longer foraging trips during incubation when shifts average six days (Harris, 1966b), compared with chick-rearing, when each parent visits the nest at least every two days (Thompson, 1987). The species was observed around North Rona and Sula Sgeir between March and August and in low densities around Shetland, Orkney and the North Sea between May and August, remaining in the North Sea into September. Sightings in the North Sea were restricted to areas off the north-east coasts of Scotland and England, and especially the Moray Firth, Firth of Forth and Flamborough Head. The Manx Shearwater is a designated feature of the Outer Firth of Forth and St Andrews Bay Complex SPAfrom the ESAS database, that were collected between 1979 and 2006, showed distributions of Manx Shearwaters during the period May–October very similar to those described above, and during the period October–November low numbers were recorded, with small concentrations in the Celtic and Irish Seas and to the south-east of Orkney (Kober et al., 2009).
Waggitt et al. (2020) produced monthly distribution maps for seabirds in the north-east Atlantic based on distribution models using data collated from aerial and vessel surveys between 1980 and 2018. These maps show similar Manx Shearwater distributions in Scottish waters to Stone et al. (1994) and Kober et al (2009), with the highest densities to the west of Scotland, and densities increasing through the breeding season, peaking in August.
Projected distributions based on foraging range and colony sizes also indicate a Manx Shearwater hotspot to the west of Scotland (Critchley et al., 2018).
4.2.1.2.2 Derived from tracking data
Manx Shearwaters have been tracked using GPS tags from breeding colonies on Rum in Scotland, Copeland in Northern Ireland, High Island and Great Blasket in south-west Ireland, Lundy in the Bristol Channel, and Skomer and Skokholm in south-west Wales (Guilford et al., 2008, Dean et al., 2010, Freeman et al., 2012, Dean et al., 2013, Dean et al., 2015, Padget et al., 2019, Critchley et al., 2020, Kane et al., 2020). Prior to laying, females undertake a "pre-laying exodus", during which they may travel a considerable distance to productive feeding grounds (e.g. at the edge of the continental shelf) which are not regularly visited when birds are attending eggs or chicks (Dean, 2012). During the breeding season birds from all colonies (except Lundy and Skokholm, where fewer birds have been tracked to date) visited Scottish waters. Birds tracked from Rum moved extensively through the North and West SMP regions (Dean et al., 2015, Padget et al., 2019), those from High Island and Great Blasket moved extensively through the West SMP region and into the North SMP region (Wischnewski et al., 2019, Kane et al., 2020), and birds from Copeland used the West SMP region extensively (Dean et al., 2015, Padget et al., 2019). The Solway and Clyde Scottish Marine Regions (SMRs) were used by birds from multiple colonies at all stages of the breeding season, although the extent of use varied between years (Dean et al., 2013, Dean et al., 2015).
In September, Manx Shearwaters depart in a south-westerly direction from the colonies to their wintering areas off South America, and arrive back from a westerly direction between March and May (Guilford et al., 2009). Fledglings depart on migration almost immediately after fledging and take a similar but more direct migration route than breeding adults (Wynn et al., 2021).
Few data exist on the distribution of immature Manx Shearwaters in Scottish waters. Immatures were tracked from Skomer in June–July 2013 and 2014 and, while adult movements extended into Scottish waters, immatures remained significantly further south, and there was little overlap between the core use areas of adults and immatures (Fayet et al., 2015).
4.2.2 Population status and abundance
4.2.2.1 Breeding
The most recent global population estimate for Manx Shearwaters is 338,000–411,000 pairs, with 305,000–374,000 of these breeding in Britain and Ireland. Scotland holds internationally important numbers of the species, with an estimated 126,545 (95% CLs 112,285–141,701) breeding pairs (Table 1), representing 38% of the population in Britain and Ireland (Mitchell et al., 2004). The breeding colony on Rum was estimated as 120,000 (107,000–134,000) breeding pairs in 2001 (Mitchell et al., 2004), making it the largest single-island colony in the world at the time. A more recent survey conducted in 2021, using slightly different survey methods, estimated the colony size as 288,894 (226,010 – 403,915) pairs (Inger et al., 2022). It is not clear to what extent the difference in survey estimate represents a genuine increase in population size since estimates of nesting density were similar in both surveys. The apparent increase in population size in 2021 resulted almost entirely from an increase in the size of the area considered to be suitable for nesting. The colonies at both Rum and St Kilda qualify as internationally important (Mitchell et al., 2004).
| Administrative area |
SPA colonies |
AOS |
95% LCL |
95% UCL |
Colonies counted |
Colonies not counted |
|---|---|---|---|---|---|---|
| Shetland |
Non-SPA |
7 |
7 |
7 |
1 |
|
| Western Isles |
St Kilda |
4,803 |
3,593 |
5,909 |
11 |
|
| Lochaber |
Rum |
120,000 |
107,000 |
134,000 |
1 |
|
| Non-SPA |
252 |
202 |
302 |
3 |
||
| Argyll & Bute |
Non-SPA |
1,483 |
1,483 |
1,483 |
2 |
1 possible |
| Cunninghame |
Non-SPA |
Present? |
0 |
1 |
||
| Kyle & Carrick |
Non-SPA |
Present? |
0 |
1 |
||
| Total |
126,545 |
112,285 |
141,701 |
8 |
3 |
1 Four islands of St Kilda treated as one colony.
4.2.2.2 Non-breeding
Skov et al. (1994) estimate that immatures could comprise around 25% of the entire population of Manx Shearwaters and Guilford et al. (2008) estimate that up to half the population could be non-breeders, in any given year (i.e. including birds of breeding age that were unpaired). An estimated 200,000 individuals are present in Scottish waters on passage (Furness and Wade, 2012).
4.2.3 Productivity and survival
4.2.3.1 Age at first breeding
Age at first breeding is not known for Manx Shearwaters breeding in Scotland but studies at the Pembrokeshire colonies suggest that some will breed from three years old, but that most do not breed before the age of five (Harris, 1966a).
4.2.3.2 Productivity
In Scotland, productivity data are collected on Rum, Canna and Sanda, and the average productivity between 1986 and 2018 was 0.62 chicks fledged per AOS (JNCC, 2021b), with no indication of a temporal trend over this period. Productivity as high as 0.97 has been recorded on Sanda (Mavor et al., 2006).
More experienced breeders may have higher productivity than first-time breeders. Hatching success on Skokholm between 1973–76 was lower in newly formed pairs (66.2%) than established pairs (79.2%; Brooke, 1978).
Manx Shearwater burrows can be susceptible to flooding and high rainfall during incubation has been shown to reduce hatching success on Rum and Canna (Thompson and Furness, 1991).
4.2.3.3 Survival
The average annual adult survival rate on Skokholm was estimated to be 0.902 between 1963 and 1968, ranging from 0.794 to 0.965 (Harris, 1966a, Perrins et al., 1973). Mean adult annual survival on Skomer was 0.87 between 1978 and 2018 (Zbijewska et al., 2020). Data from Rum (Hallival, Askival and Trollaval) from 1994–2014 and Sanda Island from 2000–2005 indicate a mean (± SD) annual adult survival rate of 0.93 ± 0.03 (Horswill et al., 2016).
Estimating survival for non-breeders is challenging due to the low re-encounter rates, but Harris (1966a) estimated that pre-breeders frequenting the Skokholm colony, from the age of four years old, have an annual survival rate of around 0.8. On Skokholm, average survival rates of young Manx Shearwater from fledging to returning to the colony as pre-breeders were estimated as 26.7% for 1964–69 (Perrins et al., 1973) and 21.8% for 1967–71 (Brooke, 1977), but these are likely to be underestimates as not all surviving birds will be recaptured, and Perrins (2014) suggests the actual survival rate is around 28–37%. The percentage of birds surviving to breed will be lower since pre-breeders spend more time on the surface at the colony and are at higher risk of predation than breeding birds (Perrins et al., 1973). There is some indication that chicks fledging later in the season have lower survival rates (Perrins, 1966).
4.2.4 Foraging ecology
The Manx Shearwater is a pursuit-plunger (Brown et al., 1978), and birds studied at Skomer had an average dive depth of 9.6 m, and a maximum depth of 55 m, with dives lasting 13.5 seconds on average and a maximum of 46 seconds, with no differences between the sexes (Shoji et al., 2016). For birds tracked from Skomer, diving occurred during the day and peaked in the evening (Shoji et al., 2016), but nocturnal foraging was observed from tracking of birds from High Island, Ireland (Kane et al., 2020). The species displays a dual foraging strategy during the breeding season, undertaking a combination of short trips for chick provisioning and long trips for self-provisioning (Shoji et al., 2015, Wischnewski et al., 2019). Since dual foraging can lead to bimodality in foraging ranges, it should be considered when using foraging range data to assess the risks posed by wind farm developments.
Currently, the standard foraging ranges for Manx Shearwater used in offshore wind casework come from Woodward et al. (2019), who collated the available data to give a maximum foraging range of 2890 km, a mean maximum of 1346.8 ± 1018.7 km, and a mean of 36.1 ± 88.7 km. Foraging trip durations and distances vary between years, colonies, breeding stage and the sexes (Thompson, 1987, Gray and Hamer, 2001, Guilford et al., 2008, Dean et al., 2010, Dean, 2012, Dean et al., 2013, Dean et al., 2015, Wischnewski et al., 2019). Published tracking data from Scotland is limited to 75 trips by 20 chick-rearing birds on Rum, which each lasted one day and had a median total trip distance of 184 km (interquartile range 128–274 km) and median maximum distance from the colony of 35 (29–73) km (Dean et al., 2015). The distances recorded by birds from Rum were shorter than those of chick-rearing birds from other colonies tracked simultaneously, the longest of which were undertaken by birds from Skomer, which covered a total of 297 (203–581) km during trips of one to two days, and reached a median maximum distance of 86 (61–134) km from the colony (Dean et al., 2015). Manx Shearwaters tend to travel further during incubation than chick-rearing, with incubating birds from Skomer undertaking trips of 8 (7–11) days duration, covering total distances of 1,517 (925–2,117) km and reaching a maximum distance from the colony of 254 (176–295) km (Dean et al., 2015). In some years, pre-laying females from Skomer have been found to make long trips to or beyond the continental slope, up to 727 km from the colony (Dean, 2012). Birds tracked from colonies in Ireland had foraging ranges of up to 1,109 km (Wischnewski et al., 2019).
Of the 528 trips from four colonies (Rum, Copeland, Skomer and Lundy) studied by Dean et al. (2015), almost all were largely restricted to the waters over the continental shelf, with only six trips extending beyond the shelf edge. Birds foraged in areas near their respective colonies, with little overlap between colonies, but individuals from all colonies also travelled to a more distant shared foraging area at the highly productive Irish Sea Front and nearby stratified waters of the Western Irish Sea (Dean et al., 2015). This productive area is approximately 375 km from Rum and was visited on 60% of the long-distance trips made from the colony (Dean et al., 2015). There is some evidence that the foraging areas of birds from the Pembrokeshire colonies may have shifted northwards since the 1950s (Guilford et al., 2008).
Manx Shearwaters gather in dense flocks on the sea in the vicinity of breeding colonies from late afternoon, before coming ashore after nightfall. This so-called "rafting" behaviour was studied using radio telemetry of chick-reading adults at Rum, Bardsey and Skomer to inform designation of colony extension marine protected areas (McSorley et al., 2008, Wilson et al., 2009). The maximum extent of rafts (95% kernel contours of raft locations) varied according to colony: 4, 6 and 9 km respectively for Skomer, Rum and Bardsey. Locations of radio-tagged individuals in rafts were estimated by triangulation from adjacent coasts, though some potential raft locations were unobservable due to the lack of direct line-of-sight from tracking locations. It would be instructive to analyse the large body of tracking data collected using GPS tags in recent years to improve our understanding of rafting behaviour. Dean et al. (2013) found that GPS tagged birds tended to roost on the sea within 20 km of the colony prior to landfall and that they resumed roosting on the sea adjacent to the colony after their visit.
4.2.4.1 Diet
Limited dietary analysis has been conducted for Manx Shearwaters. Thompson (1987) collected 104 samples on Rum in the 1980s and found that 65.8% contained fish (clupeids, sandeels and one sheppy Argentine), 47.4% squid (Ommastrephidae, Cranchiidae, Gonatidae, Onychoteuthidae and Mastigoteuthidae) and 7.9% crustaceans, although these were always associated with fish remains and were believed to be eaten by the fish rather than the shearwater directly. Most fish were likely around 5 cm or longer, and the largest intact fish was an 8 cm long sprat (Thompson, 1987). The data suggest that squid were more important before egg-laying and that the diet during chick-rearing was primarily fish, despite squid likely being over-represented due to their digestion-resistant beaks (Thompson, 1987). Ommastrephidae squid migrate to the surface at night and the other squid families identified, plus the sheppy Argentine, are all bioluminescent, suggesting that shearwaters were feeding at night prior to chick-rearing (Thompson, 1987). All diet samples observed by Brooke (1990) on Skomer contained fish, many of them clupeids around 15 cm long.
4.3 European Storm-petrel
4.3.1 Spatial and temporal distribution in Scotland
4.3.1.1 Breeding distribution
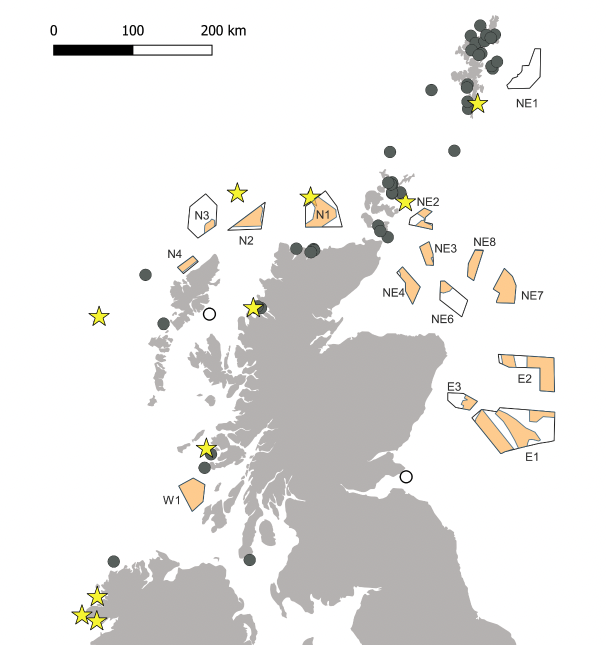
The UK and Ireland's breeding populations of European Storm-petrel are located on rat-free islands around the northern and western coasts (Figure 3; Mitchell et al., 2004, De León et al., 2006). The species is present in Scottish waters between May and October, with nests occupied between May and September, although small numbers may be present in nests in April and October (Davis, 1957a, Waggitt et al., 2020). Breeding was confirmed for the first time on the Isle of May (Firth of Forth) in 2021 (https://www.nature.scot/storm-petrels-breeding-isle-may).
4.3.1.2 Marine distribution
4.3.1.2.1 Derived from at-sea surveys
The ESAS data for 1979 to 1994 show that during May and June European Storm-petrels were mainly found over the outer shelf and shelf break to the north-west of Scotland, with low densities near Fair Isle, Shetland and the west coast of Scotland (Stone et al., 1995). In July and August the species was widespread to the north of Scotland and over the continental shelf to the west of Scotland, with densities highest at the shelf edge (Stone et al., 1995). There were also low densities around Orkney, Shetland and the north-western North Sea (Stone et al., 1995). Between September and November densities were low, but birds were observed on the continental shelf west of Scotland, along the north-east Scotland coasts, and north-east of Orkney and Shetland (Stone et al., 1995). There were no sightings in Scottish waters between December and April (Stone et al., 1995).
Poisson kriging of ESAS data collected between 1979 and 2006 showed distributions of European Storm-petrels during the breading season (June–October) broadly similar to those described above, with high densities widely distributed over the outer shelf and a large area of high density situated around 100 km north of Lewis (Kober et al. 2009).
Monthly distribution maps based on data collated from aerial and vessel surveys between 1980 and 2018 suggest a similar distribution, with European Storm-petrels present in all SMP regions and peak densities along the continental shelf, from north-east of Shetland to south-west of Ireland (Waggitt et al., 2020). Peak densities are in August, and birds are still present in October (Waggitt et al., 2020).
4.3.1.2.2 Derived from tracking data
Few tracking data exist for European Storm-petrels in the Atlantic. Bolton (2021) tracked breeding adults from the largest UK colony on Mousa, Shetland, during incubation and chick-rearing between 2014 and 2017 using GPS tags. Rather than travelling to the shelf edge, all birds used an area to the south of Shetland, moving extensively over the north-east SMP region (Bolton, 2021). A further nine chick-rearing birds GPS-tracked from Mousa in 2018 travelled in a similar direction but remained closer to the colony than birds tracked in previous years (Z. Deakin, unpublished data). The broad area of use across all years is also identified in the distribution maps produced from at-sea survey data by Waggitt et al. (2020), and broadly indicated as an area of moderate usage in Kober et al. (2009). None of the birds tracked from Mousa travelled to the west or north of Shetland, suggesting that the high concentrations of birds at the continental shelf edge are from the large colonies in the Faroes, although may also include large numbers of non-breeding birds.
Contracted by Marine Scotland, the RSPB tracked 19 breeding European Storm-petrels from Lunga, Treshnish Isles, in 2021. All birds remained on the continental shelf, moving extensively through the West SMP region, with one bird travelling 198 km from the colony to the shelf edge (RSPB unpublished data).
To date there are no published tracking studies of the migration pathways and wintering areas of European Storm-petrels breeding at Scottish colonies, although eight individuals breeding in Shetland were successfully tracked using GLS tags for 9–10 months, from the chick-rearing period in 2016 to the onset of the following breeding season in 2017 (RSPB unpublished data). A further 20 GLS tags were deployed on European Storm-petrels breeding on Lunga, Treshnish Isles in 2021 for retrieval in 2022. Limited information from ringing recoveries of birds found dead indicates the wintering areas are located off southern Africa (Marchant et al., 2002).
4.3.2 Population status and abundance
4.3.2.1 Breeding
The global population of European Storm-petrels is thought to be in the region of 1.5 million individuals (Brooke, 2004), although estimates are far from accurate and the global population trend is unknown. Data from the Seabird 2000 census suggest that Scotland holds 83% of Britain's 25,710 (95% CLs 21,043-33,517) breeding pairs of European Storm-petrels (Table 2; Mitchell et al., 2004), including the largest colony on the island of Mousa, Shetland (Mitchell et al., 2004). There were more than 50 known colonies in Scotland during Seabird 2000, but only three (Mousa, Treshnish Isles and Priest Island) were estimated to hold more than 1,000 Apparently Occupied Sites (AOS), with both Mousa and Treshnish holding internationally important numbers (Mitchell et al., 2004). Resurvey of the Mousa colony in 2008 identified a 118% increase in the population since 1996 (Bolton et al., 2010), but this growth had slowed by the most recent survey in 2015 when the population was estimated at 10,778 (95% CLs 8,857-13,207) AOS (Bolton et al., 2017). A 2018 resurvey of the Treshnish Isles of Fladda, Lunga and Sgeir a' Chaisteil estimated the population to be 8,664 AOS, representing a 109% increase since the Seabird 2000 survey in 1996 (Ward, 2018). The population at Priest Island in the Summer Isles was estimated at 3,584 ± 437 individuals in 2012, having declined by 50% since the Seabird 2000 estimate in 1999 (Insley et al., 2014), but a 2019 survey estimated 4,640 AOS, a 5% increase since Seabird 2000 (JNCC, 2021a). Resurvey of North Rona, Western Isles, in 2009 resulted in an estimate of 371 AOS, suggesting the population had remained stable since the Seabird 2000 survey in 2001 (Murray et al., 2010).
| Administrative area |
Colony |
AOS |
95% LCL |
95% UCL |
Colonies counted |
|---|---|---|---|---|---|
| Shetland Islands |
Mousa |
6,800 |
4,800 |
8,800 |
1 |
| Non-SPA |
703 |
678 |
729 |
25 |
|
| Orkney Islands |
Auskerry |
994 |
372 |
3,196 |
1 |
| Sule Skerry & Sule Stack |
309 |
309 |
309 |
1 |
|
| Non-SPA |
567 |
429 |
750 |
12 |
|
| Sutherland |
Non-SPA |
449 |
339 |
594 |
4 |
| Ross & Cromarty |
Priest Island |
4,400 |
3,300 |
6,100 |
1 |
| Non-SPA |
66 |
50 |
88 |
3 |
|
| Argyll & Bute |
Treshnish Isles |
5,040 |
5,040 |
5,040 |
1 |
| Non-SPA |
208 |
208 |
208 |
3 |
|
| Western Isles |
St Kilda |
1,121 |
825 |
2,242 |
1 |
| North Rona |
368 |
335 |
413 |
1 |
|
| Sula Sgeir |
9 |
9 |
9 |
1 |
|
| Non-SPA |
335 |
303 |
377 |
4 |
|
| Total |
21,370 |
16,997 |
28,855 |
59 |
4.3.2.2 Non-breeding
An estimated 100,000 individuals are present in Scottish waters on passage (Furness and Wade, 2012).
4.3.3 Productivity and survival
4.3.3.1 Age at first breeding
Most immatures are thought to begin prospecting at colonies in their second year and begin breeding in their third year, with almost all birds recruited into the breeding population by year five (Okill and Bolton, 2005).
4.3.3.2 Productivity
On Mousa, Shetland, breeding success from laying to fledging was 0.76 in nest boxes and 0.50 in natural nest sites in 1993; a difference that was not statistically significant (Bolton, 1996). Productivity on Skokholm in the 1950s and 60s was estimated at 0.59 and 0.49 young fledged per breeding pair by Davis (1957b) and Scott (1970), respectively. This estimate from Scott (1970) does not include data from 1967, when breeding success was only 0.27, apparently due to loss of chicks by starvation during a period of poor food availability. Mean (± SE) productivity on Skokholm for 2014–20 was 0.58 ± 0.04 (range 0.45–0.74), and was 0.80 in 2021, although the number of monitored nests is small (Brown and Eagle, 2022). Breeding success on Enez Kreiz, Brittany in 1999 was 0.53 young fledged per egg laid (Cadiou, 2001), and 0.62 at Biarritz, south-west France in 1974-79 (Hémery, 1980).
Breeding success in a population of the Mediterranean subspecies, Hydrobates pelagicus melitensis, in 1993–2006 had a mean (± SE) of 0.53 ± 0.05 but was lower in birds less than four years old (0.22 ± 0.07) (Sanz‐Aguilar et al., 2009). Higher infestations of the tick Ornithodoros maritimus have been found to reduce the body condition and survival of chicks in the Mediterranean Storm-petrel (Sanz-Aguilar et al., 2020).
4.3.3.3 Survival
Scott (1970) estimated that the annual survival rate of European Storm-petrels on Skokholm in the 1960s was 0.88-0.91. The mean survival estimate (± SD) for European Storm-petrels based on the BTO's Retrapping for Adult Survival (RAS) scheme is 0.79 ± 0.04, using data obtained without using tape-lures from Eilean Hoan, Sanda Island, Priest Island and Lunga between 1996 and 2015 (Horswill et al., 2016). A previous analysis using all BTO ring recoveries from 1967–97 estimated annual survival at 0.858 ± 0.016 (SE) (Dagys, 2001), and a similar analysis for 2001–12 suggests that survival across Britain and Ireland slowly increased during this period, staying above 0.90 from 2008–12 (Insley et al., 2014). On Priest Island, survival was 0.92 ± 0.08 (SE) in 2001–02 and 0.96 ± 0.08 in 2002–03, but was lower in 2003–12, with an overall annual survival of 0.80 during this period, and a low of 0.61 ± 0.10 in 2012 (Insley et al., 2014). Ringing data on Skomer for 2006-17 suggest an annual survival rate of 0.88 for breeding adults and 0.59 for transients (Zbijewska et al., 2020), although estimates for transient, pre-breeding birds that prospect multiple colonies before recruiting are likely to be underestimates. Survival estimates for European Storm-petrels at Aketx Islet in the Bay of Biscay for 1990–2006 ranged from 0.82–0.89, depending on the model used (Zabala et al., 2011).
Very small numbers of European Storm-petrels are ringed as chicks and immature survival rates are not currently available. Scott (1970) estimated that annual mortality between birds first returning to the colony and recruiting into the breeding population might be approximately 10-15%.
Sanz-Aguilar et al. (2008) found reduced survival in Mediterranean storm-petrels following their first breeding attempt or an unsuccessful breeding attempt.
4.3.4 Foraging ecology
European Storm-petrels are surface-feeders, although can make shallow dives up to 5 m (Flood et al., 2009, Albores‐Barajas et al., 2011), and may target areas where prey is brought to the surface by upwellings and internal waves (Scott et al., 2013). Although, for the most part, European Storm-petrels are highly pelagic during the day (Bolton, 2021), they are known to forage inshore at night and occasionally during daylight (Stegeman, 1990, Koerts, 1992, D'Elbee and Hemery, 1997, Thomas et al., 2006, Poot, 2008, Flood et al., 2009, Albores‐Barajas et al., 2011).
The continental shelf edge has been highlighted as having high concentrations of foraging European Storm-petrels (Stone et al., 1995, Waggitt et al., 2020), but breeding adults tracked from Mousa, Shetland Islands, all foraged in the shallow shelf waters to the south of the colony (Bolton, 2021). Birds from Mousa had foraging trips lasting one to three days during incubation, one day during brooding and one to two days during the post-brood stage (Bolton, 2021). Overall, the median total distance travelled was 391 km (maximum 958 km) and the median maximum range from the colony was 159 km (maximum 397 km), with no statistically significant difference between breeding stages (Bolton, 2021).
Nineteen European Storm-petrels tracked from Lunga, Treshnish Isles in 2021 (RSPB unpublished data) similarly foraged over shallow shelf waters. During the incubation stage, birds fed on average for 2–3 days at sea and ranged up to 142 km (max 198 km) from the colony, whereas when tending chicks, feeding trips tended to last for a single day and birds remained within 95 km (max 140 km) of the colony.
Six chick-rearing birds tracked from Illauntannig and High Island, Ireland, had a mean trip duration of 38 hours (maximum 67 hours), mean total distance travelled of 518 km (maximum 1,113 km) and mean range of 170 km (maximum 336) (Wilkinson, 2021). The maximum foraging range provided in the review by Woodward et al. (2019) is taken from these data alone, as presented by Critchley et al. (2018), so 336 km is the current standard foraging range used for offshore wind casework., although confidence in this value is poor. Most of the foraging locations for these birds were near or beyond the shelf edge at the Porcupine Bank and Porcupine Seabight and one bird apparently foraged close to the coast overnight (Wilkinson, 2021). There was also evidence of dual foraging, with some birds making shorter trips and remaining much closer to the colony (Wilkinson, 2021), and the possibility of birds using this strategy should be considered when using foraging range data to assess the risks posed by wind farm developments. All tracking to date in Britain and Ireland has been conducted between mid-July and mid-August, representing a fairly narrow time window relative the full extent of the breeding season (May to October).
The trips of European Storm-petrels tracked in the Atlantic are shorter than those of GPS-tracked Mediterranean storm-petrels, which lasted up to five days, covered total distances of up to 1,727 km, and ranged up to 469 km from the colony (De Pascalis et al., 2021, Rotger et al., 2021). Birds breeding at Benidorm Island in the Mediterranean had foraging areas that encompassed submarine canyons (Rotger et al., 2021). Birds breeding at Sardinia, Italy, foraged in shallow water with strong currents during their longer incubation trips but closer to the colony during chick-rearing, in shallow, productive waters (De Pascalis et al., 2021).
4.3.4.1 Diet
On Skokholm, Pembrokeshire, in the 1960s, regurgitates contained young herring Clupea harengus and sprat Clupea sprattus as well as amphipods and decapods (Scott, 1970). Two main prey groups were identified in birds breeding in the Bay of Biscay by D'Elbee and Hemery (1997). The first consisted of oceanic and neritic organisms such as ichthyoplankton (Gadidae, Ammodytidae, Myctophidae) and microzooplankton (Copepoda, Euphausiacea, Chaetognatha, Anthomedusae and meroplanktonic larvae), with an average body length of 4 cm (range 0.5–9 cm) and included bioluminescent species that migrate vertically in the water column at night. The second group consisted of littoral (Gobiidae) and suprabenthic intertidal organisms such as isopods (Cirolanidae), which were presumably collected during nocturnal inshore foraging (D'Elbee and Hemery, 1997). DNA analysis of faeces and regurgitates collected from European Storm-petrels breeding on Mousa, Shetland (Wood, 2017) identified fish in 97% of faecal samples and 92% of regurgitates, and invertebrates in 71% of faecal samples and 3% of regurgitates. No faecal samples contained squid DNA. The most common fish species were Norway pout Trisopterus esmarkii and Whiting Merlangius merlangus. European Storm-petrels will also scavenge on carcasses and waste from fishing boats and can be found foraging in high concentrations around fish farms (Hudson and Furness, 1989, Medeiros Mirra, 2010, Borg, 2012, Josa et al., 2021).
4.4 Leach's Storm-petrel
4.4.1 Spatial and temporal distribution in Scotland
4.4.1.1 Breeding distribution
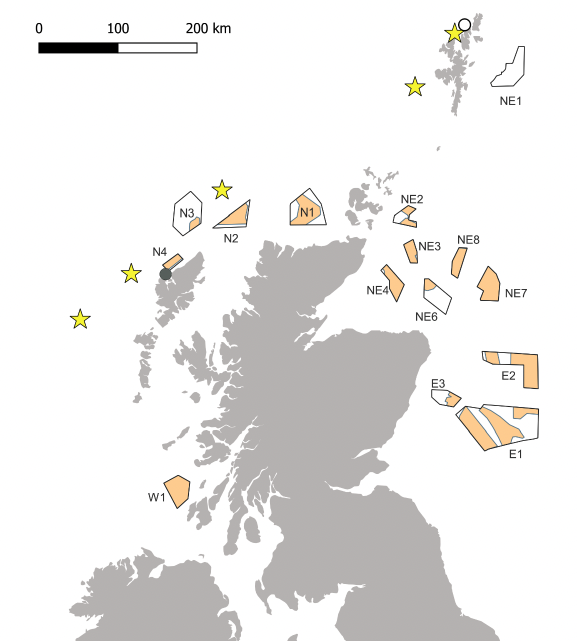
Most Leach's Storm-petrels breeding in Britain and Ireland do so at colonies in Scotland's Western Isles, with smaller numbers on islands off the west coast of Ireland, and two small colonies (± 20 pairs) in Shetland (Figure 4; Mitchell et al., 2004). However, for two of the seven Scottish colonies for which breeding was confirmed in the Seabird 2000 census, there is no recent evidence of breeding (Foula, Shetland and Old Hill, Lewis). Breeding was confirmed at Gloup Holm, Shetland in 2020 (Miles et al., 2021). Birds are present at the breeding colonies between April and October.
4.4.1.2 Marine distribution
4.4.1.2.1 Derived from at-sea surveys
Based on data from at-sea surveys, during May to August Leach's Storm-petrels were highly concentrated over the shelf edge, Rosemary Bank seamount and the deeper waters to the north-west of Scotland (Skov et al., 1994, Stone et al., 1995, Kober et al., 2009). Numbers were highest in the waters around St Kilda, with occasional sightings in the northern North Sea (Hall et al., 1987, Stone et al., 1995). The species was more widely dispersed between September and November, with low numbers around the Outer Hebrides, Shetland, and the east of Scotland (Stone et al., 1995). There were no sightings of Leach's Storm-petrels in Scottish waters between December and April.
Poisson kriging of ESAS data collected between 1979 and 2006 showed distributions of Leach's Storm-petrels during the breeding season (June – October) similar to those described above, with the highest densities on the northern slope of the Rosemary Bank seamount (Kober et al. 2009).
4.4.1.2.2 Derived from tracking data
Tracking of breeding Leach's Storm-petrels from St Kilda in 2021 confirmed their use of deep waters (>1,000 m) around the Rosemary Bank seamount that were identified as hotspots by at-sea surveys (RSPB unpublished data). There is no information from either tracking or ringing regarding the marine distribution of Leach's Storm-petrels (of any age-class) from Scottish colonies outside the breeding season. Twenty Leach's Storm-petrels breeding on St Kilda were equipped with GLS tags in 2021, for retrieval in 2022 (RSPB unpublished data).
4.4.2 Population status and abundance
4.4.2.1 Breeding
The global population of Leach's Storm-petrels is estimated at 6.7–8.3 million breeding pairs, with 40–48% of these in the Atlantic (>90% of which breed in the western Atlantic) and 52–60% in the Pacific (BirdLife International, 2022c). There is genetic evidence for long-distance dispersal between colonies in the Atlantic, suggesting that Leach's Storm-petrels in the North Atlantic should be considered as a metapopulation for conservation and management purposes (Bicknell et al., 2012). At the time of the Seabird 2000 census Scotland held more than 99% of Britain and Ireland's breeding Leach's Storm-petrels, with 94% of these on the St Kilda archipelago (Table 3; Mitchell et al., 2004). Additional surveys of Dùn, the island of the St Kilda archipelago with the largest Leach's Storm-petrel population, indicated a 54% decline between 1999 and 2006 (Newson et al., 2008). Data from a further survey of the four main St Kilda islands in 2019 suggest a 68% decline across the archipelago in the 20 years since Seabird 2000, with the population currently estimated at 8,869 (95% CLs 7,787–10,102) AOS (Deakin et al., 2021). This decline led to the species being up-listed from Amber to Red in the UK's Birds of Conservation Concern assessment (Stanbury et al., 2021). The population on North Rona was estimated at 713 AOS in 2009, a 34% decline since 2001, and a 2009 survey of Sula Sgeir found no Leach's Storm-petrels (Murray et al., 2010).
| Administrative area |
Colony |
AOS |
95% lower confidence limit |
95% upper confidence limit |
|---|---|---|---|---|
| Shetland Islands |
Foula |
15 |
3 |
30 |
| Gruney |
20 |
|||
| Orkney Islands |
Sule Skerry |
0 |
||
| Western Isles |
St Kilda |
45,433 |
34,310 |
61,398 |
| Flannan Isles |
1,425 |
1,232 |
1,708 |
|
| Loch Roag |
17 |
15 |
20 |
|
| North Rona |
1,132 |
849 |
1,700 |
|
| Sula Sgeir |
5 |
3 |
7 |
|
| Total |
48,047 |
36,432 |
64,883 |
4.4.2.2 Non-breeding
An estimated 100,000 individuals are present in Scottish waters on passage (Furness and Wade, 2012).
4.4.3 Productivity and survival
4.4.3.1 Age at first breeding
While the age of first breeding is not known for populations breeding in Scotland, in Canada Leach's Storm-petrels typically breed for the first time at 4–5 years of age (Huntington and Burtt, 1970).
4.4.3.2 Productivity
Breeding success was estimated at a minimum of 0.59 young per egg laid in burrows inspected with an endoscope on St Kilda in 2007 (Money et al., 2008) and 0.65 in 2008, with most failures occurring at the egg stage (Bicknell et al., 2009). The number of Leach's Storm-petrels breeding in nest boxes on St Kilda has increased from a single pair in 2006 to nine pairs in 2021 and breeding success has risen each year from 0.00 in 2006 to 0.89 in 2021, presumably due to increased breeding experience of nest box occupants (Nisbet, 2021). On Kent Island, New Brunswick, breeding success varied with breeding age, increasing by 0.39 ± 0.04 (SE) per year for the first two years, and declining by 0.20 ± 0.04 per year for the final two years of breeding (Mauck et al., 2012).
4.4.3.3 Survival
Our literature search found one estimate of adult survival rate (0.880) for Leach's Storm-petrel in Scotland (1984), cited in MacDonald et al. (2015, Table 1). This value was estimated in 1984 based on ringing conducted during the 1970s and early 1980s. Since that time the number of Great Skuas Stercorarius skua breeding on St Kilda has increased substantially, and they have been found to exert high levels of predation on Leach's Storm-petrels (Votier et al., 2006, Miles, 2010), with inevitable, but currently unquantified, impacts on adult survival rates. There is a need to assess current survival rates of Leach's Storm-petrels on St Kilda.
Mean annual survival was 0.78 ± 0.04 at Bon Portage Island, Nova Scotia in 2009–14 (Fife et al., 2015), and 0.79 for colonies at both Baccalieu Island and Gull Island, Newfoundland (Pollet et al., 2019), which is considered low for Procellariiformes and appears to be driving the decline of the species in the north-west Atlantic. Estimates on Kent Island, New Brunswick, Canada suggest that survival increased with age, with annual survival of 0.749 ± 0.046 from breeding year 1 to 2, 0.802 ± 0.040 from 2 to 3, and 0.870 ± 0.030 thereafter (Mauck et al., 2012). Estimates of adult survival for two breeding locations in the East Pacific (Rock Island and Cleland Island, British Columbia) were considerably higher at 0.975 ± 0.011 and 0.975 ± 0.001, respectively (Rennie et al., 2020).
4.4.4 Foraging ecology
Leach's Storm-petrels are highly pelagic, foraging over deep waters, although the species has been observed foraging on the shoreline in eastern Newfoundland, apparently as a result of extreme food stress caused by a marine heatwave (D'Entremont et al., 2021). Incubating adults tracked from Country Island and Bon Portage Island in Nova Scotia travelled to or beyond the continental shelf with foraging ranges of 1,015 ± 238 km and 612 ± 166 km, respectively (Pollet et al., 2014). Foraging range varied between years (Pollet et al., 2014). Tracking of incubating Leach's Storm-petrels at seven colonies in eastern Canada (including Country Island and Bon Portage) revealed little overlap between foraging areas among colonies, although there was no evidence that this was due to intra-specific competition rather than the availability of pelagic foraging habitat (Hedd et al., 2018). Foraging range was not affected by colony size but tended to be shorter for birds breeding at the southern end of the range (Hedd et al., 2018). Trips averaged 4.0 ± 1.4 days, with birds travelling to or beyond the continental slope to highly pelagic waters 400–830 km from the colonies, on average (Hedd et al., 2018). Birds from all but one colony showed a preference for deeper waters, with tracked birds from five of the seven colonies foraging over waters with median depths of more than 1,950 m and average chlorophyll a concentrations less than 0.6 mg/m3 (Hedd et al., 2018). In contrast, birds from the most southerly colony, Kent Island, mainly foraged in shallower neritic waters with a median depth of 181 m (Hedd et al., 2018). Woodward et al. (2019), the current standard reference for foraging ranges for UK offshore wind casework, present a mean foraging range of 657 km, which is based on 11 individuals from Gull Island, Newfoundland, which were the only birds tracked by Hedd et al. (2018) using GPS devices, rather than geolocators, which are less precise. Further GPS tracking has been carried out at Gull Island and the maximum foraging range for 182 complete trips during 2016–2021 was 953 km (Collins et al., 2022).
Fourteen Leach's Storm-petrels tracked from St Kilda in 2021 (RSPB unpublished data) foraged over deep (>1000 m) oceanic water to the north and north-west of St Kilda. During the incubation stage, birds fed on average for 2–3 days at sea and ranged up to 301 km (max 412 km) from the colony, whereas when tending chicks, feeding trips tended to last for two days and birds ranged up to 260 km (max 294 km) from the colony.
In their wintering areas off southern Africa, Leach's Storm-petrel abundance peaked in waters more than 2,000 m deep, particularly in areas with large salinity and sea surface temperature gradients (Camphuysen, 2007).
4.4.4.1 Diet
As far as we are aware there have been no studies of the diet of Leach's Storm-petrels in the UK, but analysis of 18 regurgitate samples from the Faroes found them to be comprised of fish (primarily Gadiformes),crustaceans and other invertebrates (Hey, 2019). The fish species consumed included the demersal species Norway Pout Trisopterus esmarkii and Whiting Merlangus and, in contrast to studies in the western Atlantic (see below), Hey (2019) found no pelagic fish species, suggesting that foraging habits may differ for birds breeding in the Faroes.
In Newfoundland, fish and crustaceans appear to make up the bulk of the diet of chick-rearing birds, with fish increasing through the breeding season, and mature myctophids and sandeels being particularly abundant in regurgitate samples (Hedd and Montevecchi, 2006, Hedd et al., 2009). Prey items collected range in size from 0.3 to 18 cm (Hedd et al., 2009). While myctophids are deep-water species, migrating to the surface at night, the amphipod Hyperia galba, abundant in diet samples, is a parasite of jellyfish commonly found inshore, suggesting both offshore and nearshore foraging by Leach's Storm-petrels. Other prey items include euphausiids, decapods, copepods, isopods and cephalopods (Montevecchi et al., 1992, Hedd and Montevecchi, 2006, Hedd et al., 2009). A more recent study of the diet of Leach's Storm-petrels at colonies in Nova Scotia and New Brunswick found that fish occurrence was lower and cephalopod occurrence was higher than in the Newfoundland studies, although fish still comprised the bulk of the diet (Frith et al., 2020). Diet samples from Daikoku Island, Hokkaido, Japan, consisted of similar taxa to those collected in the north-west Atlantic, although the species differed and cephalopods were more prominent (Watanuki, 1985). Leach's Storm-petrels will also scavenge on fisheries discards (Frith et al., 2020).
Leach's Storm-petrels are surface-feeders and the prominence of diel vertically migrating prey such as myctophids in their diet suggests that they feed mainly at night (Hedd and Montevecchi, 2006). However, the species has been observed feeding during daylight on the midwater fish Vinciguerria lucetia in the tropical Pacific when large numbers of fish apparently came to the surface to feed on an abundance of copepods, concentrated by an oceanographic anomaly, so care should be taken when assuming foraging habits based on diet (Pitman and Ballance, 1990).
4.5 Northern Fulmar
4.5.1 Spatial and temporal distribution in Scotland
4.5.1.1 Breeding distribution
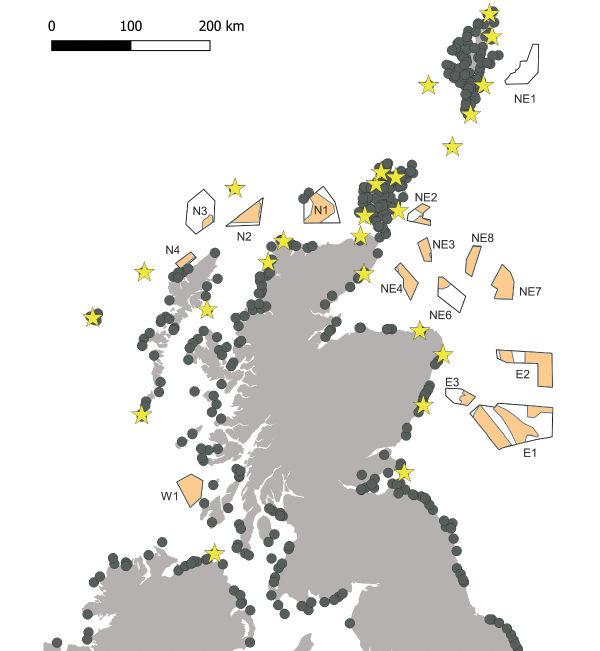
Before the mid eighteenth century, the only Northern Fulmar breeding sites within Britain and Ireland were at St Kilda but following a rapid population and range expansion between the late 19th and late 20th centuries the species now breeds all around the coasts of Britain and Ireland, both on mainland cliffs and offshore islands. The largest colonies and most breeding birds are in the north and west of Scotland (Mitchell et al., 2004).
4.5.1.2 Marine distribution
4.5.1.2.1 Derived from at-sea surveys
Fulmars breed in Scotland between April and mid-September but are present in Scottish waters throughout the year and may visit nest sites from November (Fisher, 1952). During March and April, the highest densities of Fulmars observed on at-sea surveys in Scotland were around the shelf edge to the north and west, including Shetland (Stone et al., 1995). During May to July the highest densities remained at these shelf edges, but the densities increased around colonies in Shetland, Orkney and the Hebrides. Fulmars leave the colonies during September, and from August to November high densities were found throughout the northern North Sea, as well as to the north and west of Scotland, and around Shetland and Orkney (Stone et al., 1995). The species remains in Scottish waters over winter. The results from Stone et al. (1995) are supported by those of (Kober et al., 2009) and Waggitt et al. (2020) who modelled at-sea survey data, and Darby et al. (2021), who modelled habitat preference based on tracking of 102 breeding adults from 11 colonies between 2009 and 2019.
4.5.1.2.2 Derived from tracking data
Tracking from Eynhallow revealed high variation in foraging trips during incubation, with Fulmars foraging in the North Sea, Norwegian Sea, Barents Sea and the central North Atlantic (Edwards et al., 2013, Edwards, 2015, Edwards et al., 2016), but trips during chick-rearing remained largely over the Scottish continental shelf and northern North Sea (Edwards, 2015). A male Fulmar tracked from Eynhallow to the Mid-Atlantic Ridge during incubation foraged over areas of persistent thermal fronts along the Charlie-Gibbs Fracture Zone (Edwards et al., 2013).
4.5.2 Population status and abundance
4.5.2.1 Breeding
The global population of Northern Fulmar is estimated at approximately 7 million pairs, or 20 million individuals (Carboneras et al., 2016), with 3.38–3.50 million pairs in Europe (BirdLife International, 2022b). At the time of the Seabird 2000 census, there were an estimated 537,991 AOS in Britain and Ireland, of which 90% (485,852 AOS) were in Scotland (Mitchell et al., 2004). The population growth in Britain appeared to have stopped between the 1980s and 2000, with overall numbers remaining stable but a mixture of increases and decreases at individual colonies (Mitchell et al., 2004). Scottish populations have been declining since the 1990s and in 2019 (the latest year for which data are available) were at their lowest since nationwide monitoring began in 1986 (JNCC, 2021b).
4.5.2.2 Non-breeding
An estimated 1 million Fulmar are present in Scotland outside of the breeding season (Furness and Wade, 2012).
4.5.3 Productivity and survival
4.5.3.1 Age at first breeding
The mean age at first breeding in the Northern Fulmar is 9 years (range 6–12 years; Dunnet and Ollason, 1978b), but this may be an overestimate (Dunnet et al., 1979). More recent estimates would be useful considering the change in the trajectory of Scotland's Fulmar population since the 1970s.
4.5.3.2 Productivity
In Scotland, Fulmar productivity has been declining since the mid-1990s, and the number of young fledged per pair decreased from c. 0.55 to 0.39 between 1986 and 2019 (JNCC, 2021b). Productivity is monitored annually at the Isle of May (Newell et al., 2016), Fair Isle (Shaw et al., 2002), Canna (Swann, 2000) and Eynhallow, Orkney (Lewis et al., 2009). Mavor et al. (2008) present multi-year data for a large number of colonies in Scotland.
4.5.3.3 Survival
Annual adult survival has been estimated over several decades at Eynhallow by Grosbois and Thompson (2005). Survival estimates for the first time interval of the study (1962–1963) were 0.951 (95% confidence interval = 0.911–0.973) for females and 0.975 (0.9592–0.9888) for males, but declined over time and for the last time interval (1994–1995) were 0.898 (0.843–0.936) for females and 0.8674 (0.8105–0.9091) for males. Previous studies provide estimates of adult survival at Eynhallow going back to the 1950s (Dunnet et al., 1963, Dunnet and Ollason, 1978b, Dunnet and Ollason, 1978a).
Data on the survival of juvenile and immature Fulmars are lacking. Dunnet and Ollason (1978b) used indirect methods to estimate that mean annual survival of pre-breeders in the 1950s–1970s was between 0.88 and 0.93, but this may be an overestimate.
4.5.4 Foraging ecology
Fulmars are predominantly surface-seizers but will also make shallow dives (Garthe and Furness, 2001). The species is a common scavenger at fishing vessels (Garthe and Hüppop, 1994), where it can congregate in large numbers (Camphuysen et al., 1995), but the evidence for fisheries driving the at-sea distributions of Fulmars is mixed, with some studies suggesting the spatial distribution of fisheries and Fulmar are not correlated (Camphuysen and Garthe, 1997), while others suggest a strong correlation (Darby et al., 2021).
Woodward et al. (2019), the current standard reference for foraging ranges for UK offshore wind casework, present a mean foraging range of 134.6 ± 90.1 km, a maximum range of 2,736 km, and a mean maximum range of 542.3 ± 657.9 km.
At some colonies, including St Kilda, the timing of colony attendance and/or the prevalence of nocturnally vertically migrating species such as lantern fish in the diet suggest that Fulmars are largely foraging at night (Furness and Todd, 1984, Danielsen et al., 2010, Danielsen, 2011), while elsewhere there appears to be little or no nocturnal foraging (Furness and Todd, 1984, Ojowski et al., 2001). Tracking data also indicates nocturnal foraging around fishing vessels (Dupuis et al., 2021), and Fulmars have been observed to feed on discards at night (Garthe and Hüppop, 1993).
4.5.4.1 Diet
The diet of Northern Fulmars in Scotland has been relatively well studied during the breeding season, and far more information is available than for other Procellariiformes in the region. The studies published to date reveal that Fulmars have a very broad diet, which can vary substantially between colonies and years (Furness and Todd, 1984, Camphuysen and van Franeker, 1996, Phillips et al., 1999b), as well as between the sexes and breeding stages (Ojowski et al., 2001, Owen et al., 2013).
Diets of Fulmars at Scottish colonies include fish, crustaceans, jellyfish, squid, pelagic zooplankton and offal (Furness and Todd, 1984, Fowler and Dye, 1987, Camphuysen and van Franeker, 1996, Bourne, 1997, Hamer et al., 1997, Phillips et al., 1999b). Several studies have suggested that sandeels (Ammodytidae) are particularly important in the diets of Fulmars breeding in Shetland (Furness and Todd, 1984, Hamer et al., 1997, Phillips et al., 1999b), and Gray et al. (2003) found that chick survival was reduced in a year of low sandeel abundance. In contrast, Ojowski et al. (2001) found that sandeels comprised only 1% of diet on Foula and Unst, where Gadidae, and Norway pout in particular, were the most common prey.
4.6 Sooty Shearwater
4.6.1 Spatial and temporal distribution
4.6.1.1 Breeding distribution
Sooty Shearwaters mainly breed on offshore islands around New Zealand and Chile, with smaller colonies recorded in southern Australia and the Falkland Islands (Brooke, 2004, Reyes-Arriagada et al., 2007, Catry et al., 2019, Clark et al., 2019, BirdLife International, 2022a). They are present at the breeding colonies between late September/early October and April (Richdale, 1963, Warham et al., 1982, Brooke, 2004).
4.6.1.2 Marine distribution in Scotland
The species undertakes one of the longest known avian migrations and tracking of non-breeding Sooty Shearwaters has uncovered a clockwise circular route in the Atlantic (Hedd et al., 2012, Bonnet-Lebrun et al., 2021). Those found around Scotland are likely to be birds that migrate from colonies around Cape Horn and the Falkland Islands (Phillips, 1963, Hedd et al., 2012, Catry et al., 2019). The species has been observed in Scottish waters between May and December, but mostly at low densities (Phillips, 1963, Camphuysen, 1995, Stone et al., 1995). Long-term data from vessel-based surveys show relatively low densities during July and August, with the highest concentrations in the Minch, and moderate densities around the Rockall Bank (Stone et al., 1995). The species is widespread but at mostly low densities over the continental shelf to the north and west of Scotland, around Shetland, the Moray Firth and the North Sea. During September to November, the highest densities are around Orkney and Caithness, with widespread low or moderate densities elsewhere and more widespread, low densities off the north-east coast and in the Firth of Forth than earlier in the year (Stone et al., 1995).
4.6.2 Population status and abundance
4.6.2.1 Breeding
Sooty Shearwaters are one of the most abundant shearwaters in the southern hemisphere, but the global population is thought to be in moderately rapid decline due to the impact of fisheries bycatch, climate change and the harvesting of chicks by indigenous communities, and the species is listed as Near Threatened (Newman et al., 2008, Newman et al., 2009b, BirdLife International, 2022a). The global population is thought to number around 4.4 million breeding pairs, with 19–23.6 million individuals in total (Newman et al., 2009b, Waugh et al., 2013, BirdLife International, 2022a).
4.6.2.2 Non-breeding
An estimated 7,500 Sooty Shearwaters are present in Scottish waters outside of their austral breeding season (Furness and Wade, 2012). Declines have been observed in other wintering locations (Veit et al., 1997, Oedekoven et al., 2001), but data for Scotland are lacking.
4.6.3 Productivity and survival
4.6.3.1 Age at first breeding
The mean age at first breeding of Sooty Shearwaters is 4.8 years (range 2–10 years; Fletcher et al., 2013).
4.6.3.2 Productivity
All the productivity data found in our literature search are from New Zealand colonies, where productivity is highly variable and non-synchronous. Mean breeding success across studied colonies ranges from 3% to 76% (Jones et al., 2003, Newman et al., 2009a). Hamilton (1998) found that at colonies with high levels of predation, chick survival was 0–41%, but where there was lower natural predation or predator management had been implemented, 64–100% of chicks survived to fledging age.
4.6.3.3 Survival
Adult survival is between 86–98% (Clucas et al., 2008). Fletcher et al. (2013) estimate the survival rate of juveniles in their first two years as 41–54% per year.
4.6.4 Foraging ecology
Sooty Shearwaters forage by surface-seizing and pursuit diving (Weimerskirch and Sagar, 1996). They are capable of diving deeper than other petrels (Dunphy et al., 2015), using a zigzag pattern to reduce buoyancy (Oka, 1994). Weimerskirch and Sagar (1996) give an average dive depth of 38.7 ± 20.1 m (range 2–67 m), but Shaffer et al. (2009) found that 90% of birds dived no deeper than 30 m.
4.6.4.1 Diet
Diet data for Sooty Shearwaters in Scotland are lacking, but elsewhere they are known to feed on fish, squid and crustaceans, mostly euphausiid krill and hyperiid amphipods (Brown et al., 1981, Jackson, 1988, Shiomi and Ogi, 1992, Kitson et al., 2000, Cruz et al., 2001, Petry et al., 2008). They will also follow fishing boats to feed on discards (Wahl and Heinemann, 1979, Otley et al., 2007).
4.7 Existing documented pressures
Pressures arising from human activities in Scottish seas are comprehensively assessed in the Feature Activity Sensitivity Tool (FeAST; Rogerson et al., 2021). Here, we summarise the most important pressures for the species considered in this review.
4.7.1 Climate change
Although the effects of climate change on seabird food availability, productivity and population trends are well-documented from some seabird species and locations (Arnott and Ruxton, 2002, Frederiksen et al., 2004, Frederiksen et al., 2006, Daunt and Mitchell, 2013) there is little evidence of such direct effects on the focal species of this review. Their large potential foraging areas and generalist diets argue for resilience to climate change impacts for these species. However, Scotland's breeding populations of Leach's and European Storm-petrels are predicted to be considerably reduced or extinct by the end of the 21st century, as a result of climate change (Russell et al., 2015), likely as a result of bottom-up effects on their food resources (Daunt and Mitchell, 2013, Mitchell et al., 2020).
Climate change may result in an increase in the incidence of heavy rainfall during incubation, which is negatively correlated with Manx Shearwater hatching success on Rum and Canna as a result of burrows being flooded (Thompson and Furness, 1991). Cold temperatures early in the breeding season may also delay egg laying, which could further affect productivity (Thompson, 1987). Although Manx Shearwaters are generally believed to be robust to changes in food availability due to their ability to travel long distances and consume varied prey (Mitchell et al., 2004), milder winters have been associated with reduced prey quality, later breeding, reduced adult attendance and lower peak and fledging weights of chicks at the Pembrokeshire colonies (Riou et al., 2011). Climate change may also result in an increase in soil erosion and the consequent loss of nesting habitat, or an increase the prevalence of diseases (see below).
Milder winters may also increase the survival of invasive predators such as rats, resulting in increased predator populations and a higher rate of predation on seabirds (Swann, 2000).
Climate change may result in an increase in the incidence and severity of extreme weather events, such as storms that cause large-scale wrecks of Leach's Storm-petrels in the north-east Atlantic (Wynne-Edwards, 1953, Boyd, 1954, Teixeira, 1987).
Ocean acidification has implications for calcium-based marine organisms (Orr et al., 2005). Storm-petrels produce one of the largest eggs, in relation for female body size, of any bird (Davis, 1957a), which exerts high demands on internal calcium stores for eggshell formation. Ocean acidification could affect the ability of these species to produce well-calcified eggshells.
4.7.2 Bycatch
Bradbury et al. (2017) assessed the risk to seabirds of surface, pelagic and benthic fisheries bycatch in UK waters based on (i) species-specific sensitivity (related to conservation status, life history, behavioural traits, bycatch literature and expert opinion) and (ii) the overlap in species distribution ("vulnerability") and relevant fishery activity in summer and winter ("exposure"). They concluded that Fulmars had extremely high sensitivity to bycatch at the sea surface (ranked second of 61 species considered, with a score of 90), Manx Shearwater ranked 8th (score = 66), Sooty Shearwater ranked 16th (score = 53), Leach's Storm-petrel ranked 39th (score = 31) and European Storm-petrel ranked 46th (score = 26). All species scored highly for their response to fishing activity, indicating their tendency to follow vessels. The storm-petrels scored lower than the shearwaters and Fulmar for surface entrapment risk (2 versus 4 and 5 for storm-petrels, shearwaters and Fulmar respectively), although instances of entrapment of both European and Leach's Storm-petrels in fishing nets have been recorded (Bradbury et al., 2017, Costa et al., 2020). All five species were assessed as having lower, and similar, sensitivities to bycatch in pelagic fisheries, due to lower entrapment risk. To date there has been a limited programme of seabird bycatch monitoring in UK waters, although instances of Fulmar bycatch in longline fisheries in Scotland have been recorded, suggesting several thousand individuals may be bycaught each year (Northridge et al., 2020). Currently there are no systematic data on rates of bycatch for other focal species in Scotland, or elsewhere in UK. Manx and Sooty Shearwater are also at risk of bycatch in fisheries operating in the southern hemisphere during their non-breeding and breeding periods, respectively (Uhlmann, 2003, Bugoni et al., 2008), but the level of risk will vary depending on the areas used by the birds (Bonnet-Lebrun et al., 2020).
4.7.3 Pollution
Levels of oil contamination of seabirds have fallen in recent decades both in UK waters (Schmitt, 2019) and elsewhere in the North Sea (e.g. Stienen et al., 2017) and chronic oil pollution is generally considered not to be an important driver of seabird population declines in Scotland (NatureScot, 2021). Procellariiformes are particularly vulnerable to plastic ingestion (Moser and Lee, 1992, O'Hanlon et al., 2017) and a programme of monitoring plastic ingestion in Fulmars in the North Sea has operated since 2002 (Van Franeker et al., 2021). Whether plastic ingestion leads to mortality and exerts population level effects on the focal species in the UK is currently unknown. In Canada, Leach's Storm-petrels have been found to contain some of the highest known levels of mercury contamination of any seabirds, but no deleterious effects have been identified (Pollet et al., 2017).
4.7.4 Invasive Non-Native predators
Invasive Non-Native (INN) species such as Brown Rats Rattus norvegicus,Black Rats Rattus rattus, various species of mustelid and feral cats Felis catus have been implicated in population declines and extirpation of Manx Shearwaters, Leach's and European Storm-petrels in Scotland and elsewhere in the UK. The breeding population of Manx Shearwaters on Canna declined by 99% between 1995 and 2004, with predation by Brown Rats and cats implicated as the primary cause (Swann, 2000, Patterson, 2006). Productivity on Canna averaged 0.6 in the 1980s, declining to <0.2 in the mid-1990s due to predation by Brown Rats Rattus norvegicus (Luxmoore et al., 2019), but increased following rat eradication in 2006, averaging 0.74 fledglings/pair between 2009 and 2017 (Luxmoore et al., 2019) and 0.90 between 2015 and 2019 (JNCC, 2021b). While breeding productivity has greatly increased, the Manx Shearwater population has not recovered and remains very small (Luxmoore et al., 2019), in contrast to the recoveries of Manx Shearwater populations following rat eradication on Ramsey (Bell et al., 2019) and Lundy (Booker and Price, 2014). Brown Rats were implicated in the decline of a Manx Shearwater colony on Eigg, where predation by native Eurasian Otters Lutra lutra may have also been a factor (Evans and Flower, 1967). Brown Rats are also present on Rum and, although in the 1980s it appeared that the location of the Manx Shearwater colony on a mountain 'island' surrounded by unproductive moorland offered the species some protection from rat predation (Thompson, 1987), more recently there have been concerns that the rats may be causing a problem (Mitchell et al., 2004). There is mixed evidence regarding the effect of rats on the productivity of breeding Manx Shearwaters on Rum (Thompson, 1987, Mitchell et al., 2004, Ratcliffe et al., 2009, Lambert et al., 2015) and some evidence that negative effects of Wood Mice Apodemus sylvaticus can substitute those of rats when rat numbers are reduced (Lambert et al., 2021).
Invasive mammalian predators are a key threat to both Leach's and European Storm-petrel populations, with the species breeding almost exclusively on rat-free islands, and rarely coexisting with other introduced mammalian predators (De León et al., 2006). The colonies of both species on Foula, where cats are present, were formerly more numerous but by the time of the Seabird 2000 census had apparently been restricted to a small number of inaccessible ledges (Mitchell et al., 2004). Despite an abundance of suitable habitat, European Storm-petrels had not been recorded on the Shiant Isles prior to the eradication of Black Rats in 2016 (Main et al., 2019), but calling birds were heard in 2017 and successful breeding was detected in 2018 (First Storm-petrel chick for Shiant Isles (rspb.org.uk)).
During the Seabird 2000 census an American Mink Neogale vison was found on Old Hill, Loch Roag, which threatened the survival of the small colony of Leach's Storm-petrels that existed there at the time (Mitchell et al., 2004). Preventing the colonisation of Leach's Storm-petrel breeding islands by mammalian predators is essential for their survival. The majority of Leach's Storm-petrels in the UK breed at St Kilda, where there is a high risk of invasion by mammals as the main island of Hirta is regularly visited by supply vessels and leisure boats.
4.7.5 Native predators
Native avian predators such as gulls and skuas may become problematic if populations increase or if changes in other food sources result in increased predation of seabirds (Votier et al., 2004b, Bicknell et al., 2013, Church et al., 2019).
The large decline of Leach's Storm-petrels on St Kilda has been attributed primarily to increased predation by the Great Skua population (Votier et al., 2004a, Votier et al., 2006, Miles, 2010, Deakin et al., 2021), which increased from 10 to 271 pairs between 1971 and 1997 (Phillips et al., 1999a). Great Skuas were estimated to consume approximately 14,850 Leach's Storm-petrels at St Kilda in 1996 (Phillips et al., 1999c) and 21,000 a year in 2007–2009 (Miles, 2010).
Great Skuas on St Kilda were estimated to consume 455 Manx Shearwaters in 1996 (Phillips et al., 1999c), which is roughly equivalent to 30% of the total estimated adult mortality (Mitchell et al., 2004), and 7,450 European Storm-petrels, which was more than three times the archipelago's estimated number of breeding adults in 1999/2000 (Mitchell et al., 2004).
Leach's Storm-petrel populations on North Rona and the Flannan Isles likely also face high levels of predation by Great Skuas and Great Black-backed Gulls Larus marinus (Mitchell et al., 2004). However, populations in the western Atlantic that are not subject to intense predation have also declined (Wilhelm et al., 2020) and the species may face additional important pressures away from the breeding colonies.
Storm-petrels are vulnerable to predation by a variety of owl species, including Little Owl Athene noctua (Lockley, 1947), Short-eared Owl Asio flammeus and Long-eared Owl Asio Otis (Bried, 2003). Predation by breeding owls may contribute to storm-petrel population decline or hinder attempts to establish new colonies following INN eradication, as on Ramsey, Pembrokeshire (M. Bolton pers. obs.). Whilst no owl species breed at any Scottish storm-petrel colonies, Short-eared Owls do occur as regular migrants at colonies such as St Kilda and Mousa, but are unlikely to cause population-level impacts.
Otters will predate seabirds and have been found to impact the breeding success of storm-petrels at some colonies outside of Scotland (e.g. Quinlan, 1983), but more often their impacts at the population level are not large (Bolton et al., 2017, D'Entremont et al., 2020).
The endemic subspecies of Field Mouse on St Kilda Apodemus sylvaticus hirtensis is present at some Leach's Storm-petrel sub-colonies and seabirds are known to make up part of its diet, but it is unclear whether this is through predation or scavenging (Bicknell et al., 2009, Bicknell et al., 2020).
4.7.6 Breeding habitat degradation and loss
Decline of the European Storm-petrel population on Auskerry, Orkney, has been attributed to an increase in the number of sheep on the island, which led to the trampling and destruction of 65% of the rabbit burrows (Mitchell et al., 2004) that were previously the main nesting habitat for storm-petrels (Wood, 1997). Similarly, trampling and manuring by ground-nesting seabirds at a European Storm-petrel colony in Brittany, France, resulted in changes to vegetation cover, increased erosion and eventually the collapse of the old rabbit burrows that the storm-petrels nested in, leading to a population decline (Cadiou et al., 2010).
Competition for habitat with other ground-nesters can also be a problem. An increase in the Atlantic Puffin Fratercula arctica population on Sule Skerry apparently reduced the available habitat for European Storm-petrels by displacing them from burrows (Mitchell et al., 2004). Similarly, Manx Shearwaters breeding on Mingulay, Outer Hebrides, were apparently extirpated when their burrows were taken over by an increasing Atlantic Puffin population (Elwes and Guards, 1869).
In the western Atlantic, habitat changes at Leach's Storm-petrel breeding colonies have been associated with population declines (D'Entremont et al., 2020) and the colony on North Rona appears to be confined to the walls of man-made structures due to extensive soil erosion limiting the habitat available for burrows (Mitchell et al., 2004). High densities of Soay sheep on Hirta, St Kilda render the sward over most of the island unsuitable for breeding Leach's Storm-petrels, and most birds nest in boulder crevices. In contrast, the absence of sheep on Dùn, St Kilda allows the development of a deep tussock sward and, despite its small area, Dùn supports the majority of the UK's breeding Leach's Storm-petrels.
4.7.7 Parasites, disease and natural toxins
An outbreak of H5N1 Avian Influenza in Great Skuas in 2021, affected colonies at which Manx Shearwaters, Leach's and European Storm-petrel breed (e.g. St Kilda; Banyard et al., 2022). It is not known if Procellariiformes were also infected as none have been found dead, but they may benefit from lowered predation pressure from Great Skuas in the short term. The cause of recent wrecks of seabirds (mainly auks) in the North Sea from September 2021 to January 2022 is currently unknown, though one possibility is that they may be related to toxins associated with algal blooms, which have been detected in the blood of asymptomatic shearwaters in the Mediterranean Sea (Soliño et al., 2019). It is currently unclear to what extent petrels and shearwaters may be exposed to harmful algal toxins in Scottish waters. However, if toxic algal blooms increase in range and frequency due to climate change (Gobler, 2020), they could exert complex effects on seabirds and their predator-prey dynamics.
The disease puffinosis kills approximately 4% of Manx Shearwater fledglings on Skomer and Skokholm each year but is not considered to have population-level impacts (Brooke, 1990). While puffinosis has been linked to a coronavirus (Nuttall and Harrap, 1982), the cause is still not fully understood. A fatal case of avian malaria was recently detected for the first time in a Manx Shearwater in its wintering grounds in Brazil (Vanstreels et al., 2020).
The prevalence of parasites and disease in storm-petrels has not been well-studied at Atlantic colonies, but research on the Mediterranean subspecies of the European Storm-petrel indicates slower mass gain and reduced body condition and survival in chicks with higher parasite loads (Merino et al., 1999, Sanz-Aguilar et al., 2020).
4.7.8 Disturbance from recreation
The localised impact of disturbance by human recreational activities can be significant, with European Storm-petrels breeding within 10 m of a tourist trail on Mousa having significantly lower breeding success than those nesting in less disturbed areas (Watson et al., 2014), but due to the geographic remoteness of most colonies, and inaccessibility of the nesting areas at those colonies, most Scottish breeding colonies are not exposed to recreational disturbance. The impacts of disturbance at sea have not been studied, but Manx Shearwaters may be particularly vulnerable to disturbance by leisure craft when rafting in dense groups prior to visiting colonies.
Contact
Email: ScotMER@gov.scot