Ascribing sentience to animals and case study of the evidence for sentience in cephalopods: Scottish Animal Welfare Commission report
Scottish Animal Welfare Commission report on the principles for ascribing sentience to animals and case study of the evidence for sentience in cephalopods.
Part 1: Principles for ascribing sentience to animals
Background: Why ascribe sentience?
The need to determine which species are sentient is a welfare issue, an ethical issue, and a policy issue. Sentience is a capacity by which animals can have experiences that matter to the animal (e.g. suffering), and promoting animal welfare means increasing positive experiences and reducing negative experiences. We have a moral obligation to treat sentient animals as sentient in order to prevent or reduce unnecessary suffering and human activity should consider the impact on animals who are potentially sentient. In some cases, this should be ensured by governmental policy. Indeed, modern policy and legislation designed to protect the welfare of animals in Britain and Europe are predicated on an acceptance, underpinned by scientific opinion, that many animals are sentient (Radford, 2001). Increasingly, the debate now focuses on which animals meet the relevant threshold, and how that threshold is defined and met.
Defining sentience
The SAWC defines animal sentience as an ability to have physical and emotional experiences. These experiences matter to those animals, positively or negatively. There may be a variety of experiences in different animals.
This definition means sentience is not a particular cognitive process or behaviour per se. However, sentient animals may have different cognitive and emotional capabilities, which means that they have different needs and wants. Cognitive and behavioural capacities may also facilitate evidence of sentience (e.g. demonstrating goal-directed learning).
Ethical considerations
Our ethical duty to minimise unnecessary suffering implies a requirement to determine, robustly and fairly, which animals are sentient. Avoidable ignorance, metaphysical uncertainty or excessive scepticism are not, in themselves, valid excuses for causing suffering. We should not require impossible "proof" of sentience and avoid over-simplistic categorisations that deny sentience unscientifically (e.g. unthinking presumptions that only certain taxa can be sentient).
In ascribing sentience, we should use criteria which we would want others to apply to ourselves, defining "others" in sufficiently generic ways. We should determine practically what evidence is sufficiently convincing to affect our decisions or legislation, in a sufficiently open-minded, but not overly credulous, way. The strength and weight of evidence we consider convincing should depend on multiple factors, including the potential severity of the interventions under question (e.g. if the animals are sentient, what suffering could be caused), and their benefits to sentient animals (e.g. the benefits of biomedical research to vertebrate patients). In practice, we should err on the side of caution in avoiding suffering, except where this disproportionately increases the risks of suffering for other sentient animals.
Evidence relevant for ascribing sentience
A key question is how to evidence that an animal has passed a threshold where it should be considered sentient? Many scientists consider that we can never be completely sure that an animal is or is not sentient, but only formulate our best belief through the accumulation and review of evidence. This is a critical approach that considers the strength of evidence in proportion to our confidence that an animal has a feeling. Where the evidence is stronger, we should generally be more confident. A multifactorial, rather than a linear approach has been suggested to understand interspecies variation in states of consciousness (Birch et al, 2020). An alternative approach, based on philosophical thinking, suggests that animal emotions can be directly assessed, through observation and measurement of 'whole animal expressivity' (a process of Qualitative Behavioural Assessment or QBA; Wemelsfelder, 2007). This process considers it is possible to gain a direct window on animal sentience, rather than inferring feelings from other measures.
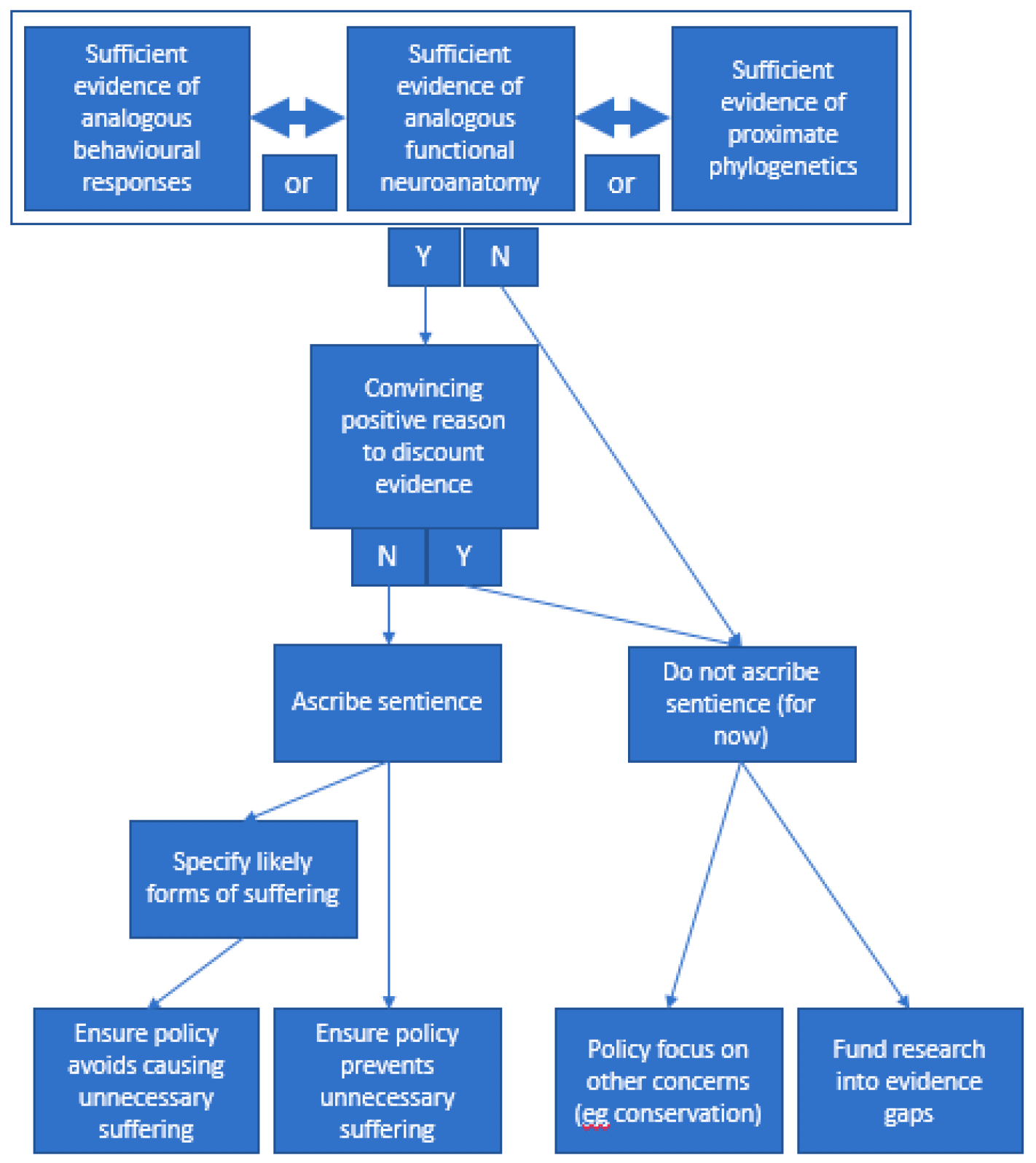
Methodologically (Figure 1), we can form plausible hypotheses of what animals may feel, and then use evidence to form conclusions. This process can be iterative, as we can only ever reach plausible but provisional views that we might change in light of further evidence. It should also aim to triangulate across all available evidence. As an overall scientific endeavour, this should inform both our ascriptions of sentience to particular animals, and the development of criteria and methods by which to do so.
Evidence of sentience might combine data from multiple sources, including:
1. Phylogenetic "proximity" to other species who have been deemed sentient
2. Neuroanatomical functioning
3. Behavioural indicators.
4. Qualitative Behavioural Assessment
1. Phylogenetic "proximity" to sentient species
One approach is to consider how closely species are phylogenetically, to humans. This assumes that the evolution of sentience occurred at some point in the phylogenetic "tree" – how far back affecting how widely to ascribe sentience (e.g. Vertebrata, Nephrozoa, Animalia). However, current evolutionary thinking suggests that complex brains and high intelligence have evolved multiple times and independently (Roth, 2015; Amodio et al., 2019). Such convergent evolution suggests that it is aspects of animals' niche or lifestyle that may be associated with the evolution of neurological or cognitive capacities (Irwin, 2020), rather than phylogenetic proximity to humans. Nonetheless, in some cases, phylogenetic proximity might be a useful practical assumption (not least in a general assumption that other humans are sentient without always assessing their individual behaviour, and also to be able to generalise conclusions from scientific evidence across a clade rather than repeating every experiment for every species or individual).
2. Neuroanatomical functioning
On the assumption that sentience can be modulated in neurological structures (e.g. brains), evidence of particular neurological functions associated with affective processing may provide support for ascribing sentience. Again, the possibility of convergent evolutions would also suggest that sentience might occur in a variety of structures (e.g. a multi-layered brain cortex; Mellor 2019), since comparable functions might be achieved by different neuroanatomical structures, so the comparison is one of analogical neurological functions rather than specific structures. It is also worth noting that the capacity for sentience per se does not obviously require the degree of anatomical complexity that is exhibited in humans.
3. Behavioural indicators
Behavioural indicators also provide evidence for valenced emotional or motivational states (on the general assumption that such states affect an animal's behaviour). Some relevant behaviours represent relatively simpler responses or motivations (e.g. withdrawal, approach, and avoidance). However, these are sometimes seen as harder to differentiate convincingly from "reflex" actions (which might theoretically occur without associated experiences). Stronger evidence may be provided by behaviours linked to processing of emotional states. For example, Sneddon (2020) suggested that sentience could be demonstrated by evidence for second order processes such as evaluating others' actions, relevant memory, and evaluative decision-making. We should remember that these may provide additional evidential support for ascribing sentience, but such higher cognitive abilities are not required for sentience (e.g. one can feel pain without being able to write a novel on it).
4. Qualitative Behavioural Assessment
Looking at animals, we do not just see physical behaviours, such as walking or eating; we also see that these behaviours are performed in different ways. There is a continuous stream of dynamic emotional expressivity in everything an animal does, indicating how it is experiencing a situation. A pig, for example, can walk towards a human in a way that is relaxed, confident, and curious, or hesitant, nervous, and on-edge. QBA is designed to describe and quantify such 'whole animal' expressivity, and has been shown, across decades of research, to be rigorous, reliable and valid in vertebrates. QBA assessments are generally found to correlate well to behavioural and physiological measurements, and to provide an additional layer of information reflecting an animal's subjective perspective that cannot be gained in other ways. This extra 'layer of information', is not merely another spoke in a wheel, but creates crucial connections between the science and practice of animal welfare, and between scientific and public views of animals
Suggested forms of evidence are listed in Table 1.
Table 1: Categories of evidence to be considered in relation to sentience
Phylogenetic proximity -
Capacity:
- Membership of humans' clade Vertebrata/Chordata/etc
- Membership of clade of another species with evidence of sentience (using other criteria)
Neuroanatomical functioning -
Capacity:- Nervous system sufficient in structure and functioning to enable experience
Example References: Winlow and Di Cosmo 2019
Mellor 2019, Sneddon et al 2014
- Possession of higher brain centre, analogous to the human cerebral cortex
Example References: Andrews et al. 2013
- Possess receptors sensitive to stimuli connected by nervous pathways to a central nervous system and brain centres
Example References: EFSA 2005, Walters 2018
Behavioural indicators -
Capacity:
- Plasticity of Behaviour
Example References: EFSA 2005
- Complexity of behaviour
Example References: EFSA 2015
- Behaviour consistent with personality
Example References: Voss 2016
- Stereotypy response to stress
Example References: BIAZA 2011
- Discrimination and generalisation
Example References: EFSA 2005
- Reversal learning
Example References: EFSA 2005
- Flexible problem solving (such as tool use)
Example References: Amodio et al 2019
- Evaluate the actions of others/form relationships
Example References: Sneddon 2020
- Integrative perceptive abilities
Example References: Macknick 2006, Hanlon et al 2009
- Spatial awareness and formation of cognitive maps, as evidence of primary consciousness
Example References: ther 2008
- Deception
Example References: Jozet-Alves et al2008
- Appropriate performance in operant studies
Example References: EFSA 2005
Specific affective states
The SAWC definition of animal sentience was one of the first to explicitly recognise that (many) sentient animals can experience a range of both positive and negative emotions. Evidence for (the capacity to have) a particular emotional state may be considered as evidence of sentience, insofar as sentience is an umbrella term for the ability to have various such states. Evidence of sentience may therefore come from evidence of (as examples): pain (see below) or malaise; discomfort; fear or distress; pleasure or enjoyment; motivation, frustration or relief; and longer-term moods.
Evidence for one state does not necessarily imply the ability for all other emotional states (e.g. an animal may be able to experience pain, and so be sentient, but not boredom). However, the SAWC assumes firstly, that if an animal can experience one such emotion, it seems more plausible that it may experience other such states in the absence of sound scientific arguments otherwise (i.e. the burden of proof shifts). Secondly, from the perspective of animal welfare, if the animal has the capacity to experience any such state, then we should consider its welfare as relevant and worthy of protection overall.
Pain and sentience
Pain is an element of sentience that merits particular consideration. Pain is the unpleasant emotional component of experiencing a noxious stimulus. It can lead to the alterations of behaviour on the expectation that sentient animals will aim to avoid or minimise pain (where possible and not outweighed by another motivation).
In addition to simple reflex responses to noxious stimuli, stronger evidence of sentience is given by demonstration that the animal has an experience of pain, for example that the animal attends to the noxious stimulus. Sneddon et al (2014) argue for two key concepts to be used to evaluate the capacity for pain:
1. Responses to noxious, potentially painful events should affect neurobiology, physiology and behaviour in a different manner to innocuous stimuli. This might be evidenced by subsequent behaviour being modified, such as avoidance learning and protective responses.
2. Animals should show a change in motivational state after experiencing a painful event such that future behavioural decision making is altered. This might be measured as a change in conditioned place preference, self-administration of analgesia, paying a cost to access analgesia or avoidance of painful stimuli and reduced performance in concurrent events
A simplified version of Table 1 can then be applied to assess whether, on the balance of evidence, an animal can be considered to be capable of experiencing pain (Table 2, adapted from Sneddon et al., 2014).
Table 2: Categories of evidence to be considered in relation to pain experiences
Neuroanatomical functioning
- Presence of nociceptors (i.e. receptors sensitive to noxious stimuli)
- Central processing in brain
- Physiological responses to noxious stimuli
- Pathways from nociceptors to CNS
- Receptors for analgesic drugs (e.g. opioid receptors)
Behavioural indicators
- Responses with high priority over other stimuli (e.g. ignores other cues)
- Behavioural changes from norm due to noxious stimuli (short and longer term)
- Altered behavioural choices/preferences (e.g. showing unwillingness to resubmit to a painful procedure
- Movement away from noxious stimuli
- Protective behaviour
- Rubbing, limping or guarding
- Trade-offs with other requirements
- Self-administration of analgesia
- No Pay cost to access analgesia
- Relief learning
- Paying a cost to avoid stimulus
- Modulation of behaviour due to analgesics
- Verbal communication of an experience associated with noxious stimuli
Evidence against sentience
We should also consider any relevant reasons to give less weight to evidence for sentience. For example, humans' programming of robots' responses means that cognitive or behavioural evidence that might legitimately support ascribing sentience to a "natural" animal should be viewed as less convincing in manmade robots. However, it is worth noting that arguments that weaken the force of evidence in implying sentience are not in themselves reasons to deny sentience.
We should avoid or try to overcome unwarranted biases towards discounting evidence in some animals that we consider convincing in others. For example, there may be cultural "prejudices" against ascribing sentience to less humanlike or familiar species (Broom 2016). For example, we should not assume that mammalian brain function is superior to other animals', especially given evidence that fish have cognitive capabilities that mammals appear to lack (Sneddon et al. 2018).
Conclusions and recommendation
The SAWC recommends that animal welfare policy in Scotland should have regard to and build on its current definition of sentience, including negative and positive welfare. This should identify which animals should be considered sentient, which specific emotions should be considered, and the implications thereof for how such animals should be treated. It is legitimate to commence the process of ascribing sentience to animals by considering the question of pain and the criteria set out above offer a useful approach.
Contact
Email: SAWC.Secretariat@gov.scot